
Провод синий желтый красный. Обозначение L и N в электрике
Переход на привычное напряжение 220 В проводился еще в годы существования Советского Союза и закончился в конце 70-х, начале 80-х. Электрические сети того времени выполнялись по двухпроводной схеме, а изоляция проводов использовалась однотонная, преимущественно белого цвета. В дальнейшем, появилась бытовая техника повышенной мощности, требующая заземления.
Схема подключения постепенно изменялась на трёхпроводную. ГОСТ 7396.1–89 стандартизировал типы силовых вилок приблизив их европейским. После распада СССР были приняты новые стандарты, основанные на требованиях Международной электротехнической комиссии. В частности, для повышения безопасности при работе в электрических сетях и упрощения монтажа, вводилась цветовая градация проводов.
Нормативная база
Основным документом, описывающим требования к монтажу электросетей, является ГОСТ Р 50462–2009, в основе которого лежит стандарт МЭК 60446:2007.
Расширенные требования к монтажу содержатся в Правилах устройства электрических установок. В них приведен рекомендуемый порядок подключения, с отсылкой к ГОСТ-Р в пунктах касающихся цветовых градаций.
Необходимость разделения по цвету
Двухпроводная система подразумевает наличие в сети фазы и нуля. Вилка для таких розеток используется плоская. Оборудование устроено таким образом, что правильность подключения роли не играет. Не важно на какой контакт будет подана фаза, аппаратура разберется самостоятельно.
При трехпроводной системе, дополнительно предусмотрено наличие заземляющей жилы. В лучшем случае, неправильное подключение проводов, приведет к постоянному срабатыванию защитного автомата, в худшем — к повреждению оборудования и пожару.
Трехпроводная система
Посмотрим на разрез трехжильного провода, который применяется для прокладки бытовых электросетей.
Цвет проводов указывает, где находятся фаза, ноль и земля. Дополнительно, на рисунке приведены типовые буквенные обозначения, применяемые в электрических схемах. Взяв в руки такой чертеж, можно визуально определить правильность выполненного подключения.
Давайте заглянем в ГОСТ и посмотрим, насколько приведенная на рисунке цветовая маркировка проводов соответствует требованиям. Пункт 5.1 общих положений содержит описание двенадцати цветов, которые должны использоваться для маркировки.
Девять цветов выделяется для обозначения фазных проводов, один для нулевого и два для заземления. Стандартом предусматривается выполнение заземляющего провода в комбинированном желто-зеленом исполнении. Разрешается продольное и поперечное нанесение полос, при это преимущественный цвет не должен занимать более 70 % площади оплетки. Отдельное использование желтого или зеленого цвета в защитном покрытии прямо запрещается пунктом 5.2.1.
Разрешается продольное и поперечное нанесение полос, при это преимущественный цвет не должен занимать более 70 % площади оплетки. Отдельное использование желтого или зеленого цвета в защитном покрытии прямо запрещается пунктом 5.2.1.
Указанная схема применяется при однофазном подключении, подходящем для большинства электрических приборов. Запутаться в ней, при правильно маркированном проводе, практически невозможно.
Пятипроводная система
Для трехфазного подключения используются пятижильные провода. Соответственно три провода выделяются под фазы, один под нейтральный или нулевой и один под защитный, заземляющий. Цветовая маркировка, как в любой сети переменного тока применяется аналогичная, в соответствии с требованиями ГОСТ.
В этом случае важным моментом будет правильное подключение фазных проводников. Как видно на рисунке, защитный провод выполнен в желто-зеленой оплетке, а нулевой — в синей. Для фаз использованы разрешенные оттенки.
С помощью пятижильных проводов можно выполнять подключение сети 380 В с правильно выполненным расключением.
Совмещенные провода
В целях удешевления производства и упрощения подключений применяются также провода двух или четырехжильные, в которых защитная жила совмещена с нейтральной. В документации они обозначаются аббревиатурой PEN. Как вы догадались, складывается она из буквенных обозначений нулевого (N) и заземляющего (PE) проводов.
ГОСТом предусмотрена для них специальная цветовая маркировка. По длине они окрашиваются в цвета заземляющей жилы, то есть в желто-зеленый. Концы должны быть в обязательном порядке окрашены в синий цвет, им же дополнительно обозначаются все места соединений.
Поскольку места, в которых выполняется подключение заранее определить невозможно, в этих точках провода PEN выделяют с помощью изолирующей ленты или кембриков синего цвета.
Нестандартные провода и маркировка
Приобретая новый провод, вы разумеется обратите внимание на цветовую маркировку жил и выберете тот вариант, где она нанесена правильно. Что делать в том случае, когда проводка уже выполнена, а цвета проводов не соответствуют требованиям ГОСТа? Выход в этом случае такой же, как и с проводами PEN. Придется выполнить ручную маркировку, после того, как вы определитесь с ролью, выполняемой подходящими к оборудованию жилами. Простым вариантом будет использование цветной изоленты соответствующих оттенков. Как минимум, стоит обозначить защитный и нейтральный провода.
Придется выполнить ручную маркировку, после того, как вы определитесь с ролью, выполняемой подходящими к оборудованию жилами. Простым вариантом будет использование цветной изоленты соответствующих оттенков. Как минимум, стоит обозначить защитный и нейтральный провода.
При профессиональном монтаже возможно применение специальных кембриков, представляющих собой полые отрезки изоляционного материала. Делятся они на обычные и термоусадочные. Вторые не требуют подбора по диаметру, но не имеют возможности повторного использования.
Встречаются также специально изготовленные маркеры, с международным буквенно-цифровым обозначением. Их применяют на вводных и распределительных щитах, к примеру, в многоквартирных домах или административных зданиях.
Цифровые метки, совместно с цветом провода, позволяют определить к какому потребителю подается питание.
Дополнительные требования
Поскольку линии, как и разводка, могут выполнятся с применением различной кабельной продукции, существует ряд правил по их взаимному подключению. Подключение трехпроводного кабеля к пятипроводному должно выполняться с соблюдением цветовой маркировки от ведущего к ведомому. Соответственно заземляющий и нейтральный цвета должны совпадать.
Подключение трехпроводного кабеля к пятипроводному должно выполняться с соблюдением цветовой маркировки от ведущего к ведомому. Соответственно заземляющий и нейтральный цвета должны совпадать.
Фазное подключение, в данном случае выполняется с использованием объединяющей шины. С одной стороны, к ней присоединяются три жилы, с другой стороны — одна, которая и будет фазой в новом ответвлении.
При монтаже бытовых электросетей, по требованиям безопасности, запрещается использовать проводку с алюминиевыми, а также многопроводными жилами. Должен использоваться только кабель с цельной медной жилой.
Трехпроводная система постоянного тока
В системах постоянного тока, также используется трехпроводная система, но назначение проводов другое. Разделение выполняется на плюсовой, минусовой и защитный. Согласно ГОСТ в таких сетях применяется следующая цветовая маркировка:
- Плюсовой — коричневый;
- Минусовой — серый;
- Нулевой — синий.
Поскольку отдельно провода под системы постоянного тока выпускать нерационально, указанная цветовая градация применяется в основном для окраски токопроводящих шин.

В заключение
Как видите, цвета проводов в электрике не прихоть производителя, а мера, направленная на обеспечение требований безопасности. При соблюдении правил монтажа обслуживать такие сети намного проще, а разобраться в подключении может не только специалист электрик, но и мы с вами.
Видео по теме
Каждый раз, когда я устанавливаю розетку или подключаю какой-то стационарный прибор встаёт вопрос о том, что значит цвет провода — фаза? Или это земля? Неразберихи добавляет то, что далеко не все кабеля — это наши родные ВВГ-3 с белым, синим и желто-зелёным проводами. Есть и китайцы с комбинациями серый + коричневый + белый, есть и сложные многожильные кабели, с которыми можно разобраться только по справочнику электрика.
В быту все эти кодировки взять неоткуда, поэтому будем ориентироваться на самую простую проводку. Простая — это кабель из трёх жил и бытовая задача, к примеру, установки розетки.
Стандартный бытовой провод с белым, синим и жёлто-зелёным цветом
Кодировка, маркировка и история
Идея разделить провода по цветам не нова — первые же эксперименты, как рисуют нам старые учебники, проводились с разноцветными клеммами и проводами. Всё та же незамутнённая простота осталась в автомобилях — синий и красный провод вряд ли перепутаешь. Правда, он иногда бывает чёрным, но это совсем другая история.
Всё та же незамутнённая простота осталась в автомобилях — синий и красный провод вряд ли перепутаешь. Правда, он иногда бывает чёрным, но это совсем другая история.
При изучении проводки самые важные для определения по цвету провода — не фаза, а земля и ноль, фазу всегда можно найти с помощью детекторной отвёртки или (практически) любого диода. А вот перепутать цвета земли и ноля иногда становится просто опасно, и определять, какого цвета провода фаза ноль земля надо заранее.
Цвет провода фазы
Как ранее было указано, особо фазу по цвету определять не требуется — почти всегда есть доступ к тому или иному инструменту для определения. Некоторый «зоопарк» в цветах наблюдается из-за того, что есть расширенные, не бытовые стандарты по цветовой дифференциации проводов, их используют настоящие электрики. Например, коричневый цвет говорит, что провод предназначен для розеток, а красный — для освещения. От этого зависит нагрузка и допустимые параметры работы.
Цвет провода земли
Заземление самый безальтернативный провод, у него всегда жёлто-зелёный цвет. Бывают отклонения, например, чисто жёлтый — когда провод импортный. В сети пишут, что встречается жёлто-зелёно-синий цвет провода, которым обозначают совмещённый рабочий нуль и землю.
Бывают отклонения, например, чисто жёлтый — когда провод импортный. В сети пишут, что встречается жёлто-зелёно-синий цвет провода, которым обозначают совмещённый рабочий нуль и землю.
Цвет провода ноля
У минуса небольшой выбор цветов — обычно это синий провод, который есть практически в любом кабеле, либо (очень редко) красный/вишнёвый. Как было сказано о земле — путать эти провода строго не рекомендуется.
Заключение
Фиксируем общую цветовую схему:
- Земля — цвет провода жёлто-зелёный или жёлтый цвет провода;
- Ноль — синий цвет;
- Фаза — цвет провода белый, красный, коричневый и любые другие незнакомые.
RozetkaOnline.ru — Электрика дома: статьи, обзоры, инструкции!
Обозначение L и N в электрике
Каждый раз, пытаясь подключить люстру или бра, датчик освещенности или движения, варочную панель или вытяжной вентилятор, терморегулятор теплого пола или блок питания светодиодной ленты, а также любое другое электрооборудование, вы можете увидеть следующие маркировки возле клемм подключения – L и N.
Давайте разберемся, о чем говорят обозначения L и N в электрике.
Как вы, наверное, сами догадались это не просто произвольные символы, каждый из них несет конкретное значение и выполняет роль подсказки, для правильного подключения электроприбора к сети.
Обозначение L в электрике
« L » — Эта маркировка пришла в электрику из английского языка, и образована она от первой буквы слова «Line» (линия) – общепринятого названия фазного провода. Также, если вам удобнее, можно ориентироваться на такие понятия английских слов как Lead (подводящий провод, жила) или Live (под напряжением).
Соответственно обозначением L маркируются зажимы и контактные соединения, предназначенные для подключения фазного провода. В трехфазной сети, буквенно-цифровая идентификация (маркировка) фазных проводников «L1», «L2» и «L3».
По современным стандартам (ГОСТ Р 50462-2009 (МЭК 60446:2007 ), действующим в России, цвета фазных проводов – коричневый или черный. Но зачастую, может встречаться белый, розовый, серый или провод любого другого цвета, кроме синего, бело-синего, голубого, бело-голубого или желто-зеленого.
Но зачастую, может встречаться белый, розовый, серый или провод любого другого цвета, кроме синего, бело-синего, голубого, бело-голубого или желто-зеленого.
Обозначение N в электрике
«N» — маркировка, образованная от первой буквы слова Neutral (нейтральный) – общепринятое название нулевого рабочего проводника, в России называемого чаще просто нулевым проводником или коротко Ноль (Нуль). В связи с этим, удачно подходит английское слово Null (нулевой), можно ориентироваться на него.
Обозначением N в электрике маркируются зажимы и контактные соединения для подключения нулевого рабочего проводника/нулевого провода. При этом это правило действует как в однофазной, так и трехфазной сети.
Цвета провода, которыми маркируется нулевой провод (нуль, ноль, нулевой рабочий проводник) строго синий (голубой) или бело-синий (бело-голубой).
Обозначение Заземления
Если уж мы говорим об обозначениях L и N в электрике, нельзя не отметить еще вот такой знак — , который также, практически всегда можно увидеть совместно с этими двумя маркировками. Таким значком отмечены зажимы, клеммы или контактные соединения для подключения провода защитного заземления (PE
– Protective Earthing), он же нулевой защитный проводник, заземление, земля.
Таким значком отмечены зажимы, клеммы или контактные соединения для подключения провода защитного заземления (PE
– Protective Earthing), он же нулевой защитный проводник, заземление, земля.
Общепринятая цветовая маркировка нулевого защитного провода – желто-зеленый. Эти два цвета зарезервированы только для заземляющих проводов и не встречаются при обозначении фазных или нулевых.
К сожалению, нередко, электропроводка в наших квартирах и домах выполнена с несоблюдением всех строгих стандартов и правил цветовой и буквенно-цифровой маркировки для электрики. И знать предназначение маркировок L и N у электрооборудования, порой, недостаточно, для правильного подключения. Поэтому, обязательно прочитайте нашу статью «Как определить фазу, ноль и заземление самому, подручными средствами? », если у вас есть какие-то сомнения, этот материал будет как нельзя кстати.
Вступай в нашу группу вконтакте!
http://rozetkaonline.ru
Как подключить вытяжной вентилятор
При подключение вытяжного вентилятора необходимо соблюдать несколько правил, которые не лишним будет повторить:
- прочитать паспорт вентилятора
- проводить подключение только при снятом напряжении сети.
- стационарная проводка должна быть оборудована автоматическим выключателем. Подключение необходимо осуществлять через автоматический выключатель QF, встроенный в стационарную проводку. Зазор между контактами автоматического выключателя на всех полюсах должен быть не менее 3 мм.
Подключение 220 В производится к вентилятору через встроенную клеммную коробку. Находится она под лицевой крышкой вентилятора. Все клеммы на вентиляторе подписаны.
- L — сюда подключаем провод с фазой,
- LT — подключаем провод с фазой, который идет от внешнего выключателя (для моделей с таймером, датчиком влажности
- N — ноль
Подключение вентилятора без датчиков
Если вам нужно подключить вентилятор без датчиков, то эта схема ваша. Любой клавишный стандартный выключатель подходит для такой задачи. В качестве включателя можно использовать регулятор скорости РС-1-300, который встраивается в стандартную монтажную коробку.
Такой регулятор скорости может включать вентилятор и менять производительность вытяжки. При уменьшении производительности уменьшается уровень шума, при необходимости быстрого удаления воздуха можно выбрать максимальную производительность.
Вентилятор с шнурком-выключателем
Включение и выключение вентилятора производится с помощью шнурочка, который прикреплен к корпусу вентилятора. К вентилятору подводится фаза и ноль.
Такие вентиляторы чаще всего используются для вентиляции кухни или других помещений, где уже заведено питание к месту установки без возможности вывести отдельный выключатель.
Вентилятор с таймером или с датчиком влажности
Схема подключения вентилятора, оборудованного таймером и вентилятора с датчиком влажности одинакова. Вентилятор с таймером начинают свою работу при подаче напряжения от внешнего включателя на клемму LT. После нажатия на клавишу выключателя, вентилятор продолжит работать время, которое выставляется на таймере. Такой вентилятор можно завязать на включение вместе с освещением.
Такой вентилятор можно завязать на включение вместе с освещением.
Такой вентилятор целесообразно использовать для вытяжной вентиляции туалета, ванной.
Вентилятор с датчиком влажности можно включить принудительно или он включается автоматически при превышении выставленного уровня влажности. Влажность устанавливается в диапазоне от 60 до 90%. Вентилятор будет работать пока влажность не уменьшится до необходимого уровня, после чего проработает еще время по таймеру задержки и выключится.
Такие вентиляторы используют для вентиляции ванной, постирочной или помещения сушки белья. Также их применяют для вытяжной вентиляции подвальных помещений частного дома с повышенным уровнем влажности.
Вентилятор с датчиком движения
Вентилятор включается автоматически при движении человека на расстоянии 1-4 метра с углом обзора 100. После того, как человека покинул помещение, вентилятор продолжит работать время по таймеру и выключится. Время задержки можно выставить под лицевой панелью вентилятора от 2 до 30 минут.
Как и вентилятор со шнурком-выключателем, такой вентилятор можно установить для вентиляции помещений, где к месту установки вентилятора выведена постоянная фаза и ноль и нет возможности установить отдельный внешний выключатель.
Подключение вентилятора оборудованного встроенной лампой
В нашем магазине есть вентиляторы, оборудованные подсветкой или лампой. Такие вентиляторы могут включаться вместе с подсветкой или вентилятор может включаться отдельно, а встроенная подсветка отдельно.
Для одновременного включения вентилятора с встроенным освещением подойдет эта схема
Для раздельного включения вентилятора и встроенного света подойдет эта схема
Конденсаторы. кодовая маркировка
Цветовая маркировка фазы, нуля и земли
Для разводки и монтажа электросетей на бытовых и на промышленных объектах, используют многожильные кабели, каждый провод внутри которых окрашен в отличительный цвет. Это необходимо, как уже было сказано, для упрощения монтажа и обслуживания сети.
Так, к примеру, если ремонт сети будет проводить человек, который не занимался её прокладкой, по цвету провода, подключенного к приборам и источникам питания, он сразу поймёт рабочую схему. В противном случае возникнет необходимость пробивать ноль и фазу вручную, используя пробник. Этот процесс непрост даже при проверке новых проводов, а при необходимости ремонта старой проводки и вовсе превратится в испытание, поскольку раньше, в советское время, маркировка проводов не осуществлялась, и все они были покрыты черной или белой изоляционной оболочкой.
Согласно разработанным стандартам (ГОСТ Р 50462) и правилам электротехнического монтажа, каждый провод, находящийся в кабеле, будь то ноль, фаза или земля, должен иметь свой цвет, который говорит о его назначении. Одним из главных требований электротехнических установок является возможность быстро и точно определить функцию провода на любом его участке. Лучше всего для решения этой задачи подходит именно цветовая маркировка.
Представленная ниже маркировка проводов разработана для сетей и электроустановок переменного тока (трансформаторы, подстанции и т. п.) с глухозаземлённой нейтралью и номинальным напряжением не более 1 кВ. Этим условиям соответствует большая часть жилых и административных зданий.
п.) с глухозаземлённой нейтралью и номинальным напряжением не более 1 кВ. Этим условиям соответствует большая часть жилых и административных зданий.
Защитный и рабочий нулевой проводник
Ноль или нейтраль на электротехнических схемах обозначается буквой N и окрашивается на всем протяжении в голубой или синий цвет без дополнительных цветовых обозначений.
PE – защитный нулевой контакт или просто «земля», имеет характерную окраску из чередующихся вдоль провода линий зеленого и желтого цвета. Некоторые производители окрашивают ее в однородный желто-зеленый оттенок по всей длине, но принятый в 2011 году ГОСТ Р 50462-2009 запрещает обозначать заземление желтым или зеленым цветом по отдельности. В сочетании зеленый/желтый эти цвета могут использоваться только в ситуации, когда обозначают заземление.
У PEN-проводов, используемых в устаревших на сегодня системах TN-C, где «земля» и ноль совмещены, более сложная маркировка. Согласно последним утвержденным стандартам, основная часть провода на всем протяжении должна быть окрашена в синий цвет, а концы и места соединения – желто-зелеными полосками. Возможно также применение проводов с противоположной маркировкой – провод желто-зеленого цвета с синими концами. Встретить такой провод в зданиях современной постройки можно редко, так как от использования TN-C отказались ввиду риска поражения людей током.
Возможно также применение проводов с противоположной маркировкой – провод желто-зеленого цвета с синими концами. Встретить такой провод в зданиях современной постройки можно редко, так как от использования TN-C отказались ввиду риска поражения людей током.
- ноль (нулевой рабочий контакт) (N) – провод синего или голубого цвета;
- земля (нулевой заземляющий) (PE) – желто-зеленый;
- совмещенный провод (PEN) – желто-зеленый с синими метками по концам.
Фазные провода
В конструкции кабелей может встречаться несколько токоведущих фазных проводов. Правилами электротехнических установок требуется, чтобы каждая фаза была обозначена отдельно, поэтому для них принято использовать черный, красный, серый, белый, коричневый, оранжевый, фиолетовый, розовый и бирюзовый цвета.
Когда проводится монтаж однофазной цепи, подключенной к трехфазной электросети, необходимо чтобы цвет фазы ответвления точно соответствовал цвету фазного контакта питающей сети, к которому она подсоединена.
Кроме того, стандартом предписывается соблюдать цветовую уникальность всех используемых проводов, поэтому фаза не может иметь такой же цвет, как ноль или земля. Для кабелей без цветовой идентификации маркировка должна быть проставлена вручную — цветной изоляционной лентой или кембриками.
Чтобы не столкнуться с необходимостью покупки термоусадочных трубок или изоленты уже во время монтажа (и не усложнить схемы лишними обозначениями), следует определиться с тем, какая комбинация цветов будет использована во всех электрических цепях дома, и закупить нужное количество кабелей каждого цвета до начала работ.
Фазный проводник, его определение по цвету или иначе
Фаза всегда монтируется проводами, изоляция которых окрашена в любые цвета, но не синий или желтый с зеленым: только зеленый или только желтый. Фазный проводник всегда соединяется с контактами коммутаторов. Если при монтаже в наличии розетки, в которых есть клемма, маркированная буквой L, она соединяется с проводником в изоляции черного цвета. Но бывает так, что монтаж выполнен без учета цветовой маркировки проводников фазы, нуля и заземления.
Но бывает так, что монтаж выполнен без учета цветовой маркировки проводников фазы, нуля и заземления.
В таком случае для выяснения принадлежности проводников потребуется индикаторная отвертка и тестер (мультиметр). По свечению индикатора отвертки, которой прикасаются к токопроводящей жиле, определяется фазный провод — индикатор светится. Прикосновение к жиле заземления или зануления не вызывает свечение индикаторной отвертки. Чтобы правильно определить зануление и заземление, надо измерить напряжение, используя мультиметр. Показания мультиметра, щупы которого присоединены к жилам фазного и нулевого провода, будут больше, чем в случае прикосновения щупами к жилам фазного провода и заземления.
Поскольку фазный провод перед этим однозначно определяется индикаторной отверткой, мультиметр позволяет завершить правильное определение назначения всех трех проводников.
Буквенные обозначения, нанесенные на изоляцию проводов, не имеют отношения к назначению провода. Основные буквенные обозначения, которые присутствуют на проводах, а также их содержание, показаны ниже.
Принятые в нашей стране цвета для указания назначения проводов могут отличаться от аналогичных цветов изоляции проводов других стран. Такие же цвета проводов используются в
- Беларуси,
- Гонконге,
- Казахстане,
- Сингапуре,
- Украине.
Более полное представление о цветовом обозначении проводов в разных странах дает изображение, показанное далее.
Цветовые обозначения проводов в разных странах
В нашей стране цветовая маркировка L, N в электрике задается стандартом ГОСТ Р 50462 – 2009. Буквы L и N наносятся либо непосредственно на клеммы, либо на корпус оборудования вблизи клемм, например так, как показано на изображении ниже.
Этими буквами обозначают по-английски нейтраль (N), и линию (L — «line»). Это означает «фаза» на английском языке. Но поскольку одно слово может принимать разные значения в зависимости от смысла предложения, для буквы L можно применить такие понятия, как жила (lead) или «под напряжением» (live). А N по-английски можно трактовать как №null» — ноль. Т.е. на схемах или приборах эта буква означает зануление. Следовательно, эти две буквы — не что иное как обозначения фазы и нуля по-английски.
А N по-английски можно трактовать как №null» — ноль. Т.е. на схемах или приборах эта буква означает зануление. Следовательно, эти две буквы — не что иное как обозначения фазы и нуля по-английски.
Также из английского языка взято обозначение проводников PE (protective earth) — защитное заземление (т.е. земля). Эти буквенные обозначения можно встретить как на импортном оборудовании, маркировка которого выполнена латиницей, так и в его документации, где обозначение фазы и нулевого провода сделано по-английски. Российские стандарты также предписывают использование этих буквенных обозначений.
Поскольку в промышленности существуют еще и электрические сети, и цепи постоянного тока, для них также актуально цветовое обозначение проводников. Действующие стандарты предписывают шинам со знаком плюс, как и всем прочим проводникам и жилам кабелей положительного потенциала, красный цвет. Минус обозначается синим цветом. В результате такой окраски сразу хорошо заметно, где какой потенциал.
Чтобы читателям запомнились цветовые и буквенные обозначения, в заключение еще раз перечислим их вместе:
фаза обозначается буквой L и не может быть по цвету желтой, зеленой или синей.
В занулении N, заземлении PE и совмещенном проводнике PEN используются желтый, зеленый и синий цвета.
На постоянном токе для проводников и шин применяются красный и синий цвета.
Цвета шин и проводов на постоянном токе
Не будет лишним показать цветовое обозначение шин и проводов для трех фаз:
Библия электрика ПУЭ (Правила устройства электроустановок) гласит: электропроводка по всей длине должна обеспечить возможность легко распознавать изоляцию по ее расцветке.
В домашней электросети, как правило, прокладывают трехжильный проводник, каждая жила имеет неповторимую расцветку.
- Рабочий нуль (N) – синего цвета, иногда красный.
- Нулевой защитный проводник (PE) – желто-зеленого цвета.
- Фаза (L) – может быть белой, черной, коричневой.

В некоторых европейских странах существуют неизменные стандарты в расцветке проводов по фазе. Силовой для розеток – коричневая, для освещения — красный.
Цветовые обозначения фазы L, нуля N и заземления
Любой электрический кабель для удобства монтажа изготавливается с разноцветной изоляцией на жилах. При монтаже стандартной электропроводки обычно используются трехжильные кабели (фаза, ноль, заземление).
Любой электрический кабель для удобства монтажа изготавливается с разноцветной изоляцией на жилах. При монтаже стандартной электропроводки обычно используются трехжильные кабели (фаза, ноль, заземление).
Фаза («L», «Line»)Основным проводом в кабеле всегда является фаза. Само по себе слово «фаза» означает «провод под напряжением», «активный провод» и «линия». Чаще всего он бывает строго определенных цветов. В распределительном щитке фазовый провод, перед тем как идти к потребителю, подключается через устройство защитного отключения (УЗО, предохранитель), в нем происходит коммутация фазы
Внимание! С голой фазой шутки плохи, по этому, чтобы не спутать фазу с чем-либо еще — запомните: контакты фазы всегда маркируются латинским символом «L», а провод фазы бывает красным, коричневым, белым или черным. Если же вы не уверены в этом или проводка устроена иначе, то приобретите отвертку с простым индикатором фазы
Если же вы не уверены в этом или проводка устроена иначе, то приобретите отвертку с простым индикатором фазы
Прикоснувшись его жалом к голому проводнику, всегда можно узнать — фаза это или нет по характерному свечению индикатора. А лучше сразу обратитесь к квалифицированному специалисту.
Ноль («N», «Neutre», «Neutral», «Нейтраль» «Нуль»)Вторым немаловажным проводом является ноль, известный в народе как «провод без тока», «пассивный провод» и «нейтраль». Он бывает только синим. В квартирных распределительных щитках его нужно подключать к нулевой шине, она помечена символом «N». К розетке провод нуля подключается к контактам, также обозначенным знаком «N».
Заземление («G», «T», «Terre» «Ground», «gnd» и «Земля»)Изоляция заземляющего провода бывает только желтого цвета с зеленой полоской. В распределительном щитке он подключается к шине заземления, к дверце и корпусу щитка. В розетках заземление подключается к контактам, обозначенным латинским символом «G» или с знаком в виде перевернутой и коротко подчеркнутой буквой «Т»
Обычно заземлительные контакты на виду и могут выступать из розеток, становясь доступными детям, что порой вызывает у многих родителей шок, тем не менее эти контакты не опасны, хотя совать пальцы туда все же не рекомендуется. Внимание! При работе с электрическими сетями под напряжением всегда велика вероятность поражения человека электрическим током или пожара
Внимание! При работе с электрическими сетями под напряжением всегда велика вероятность поражения человека электрическим током или пожара
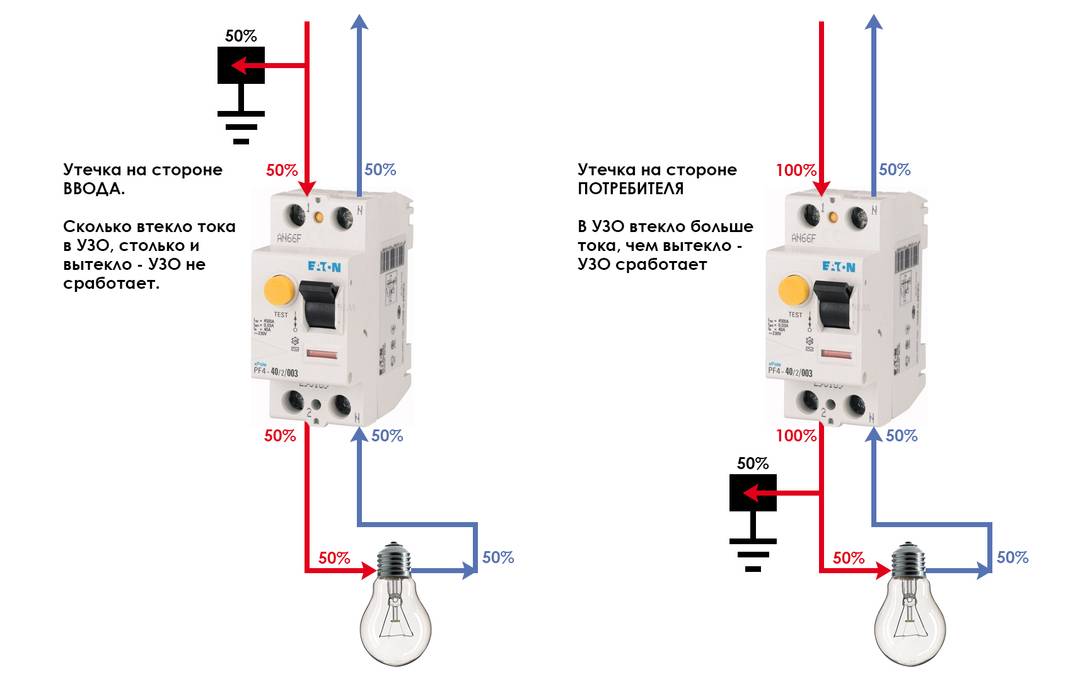
Если даже установлено УЗО, настоятельно рекомендуется соблюдать все меры предосторожности! Известно, что специальная конструкция такого выключателя сверяет синхронность работы фазы и нуля, и в случае, если УЗО обнаружит утечку тока фазы без возвращения каких-то его процентов по нулю, то немедленно разорвет контакт, что спасет человеку жизнь; однако если прикоснуться не только к фазе, но еще и к нулю — то УЗО не спасет. Прикосновение к обоим проводам смертельно опасно
Прикосновение к обоим проводам смертельно опасно.
В сетях постоянного тока
Несмотря на то что в большинстве случаев мы имеем дело с переменным током, электросети постоянного тока тоже имеют широкую сферу применения:
- В промышленной и строительной сфере – для работы электрических кранов, тележек и складского погрузочного оборудования.
- Для питания электротранспорта: троллейбусов, трамваев, электровозов, теплоходов, и т.
 д.).
д.). - Для подачи нагрузки на оперативные защитные цепи и автоматическое оборудование электрических подстанций.
Как нам известно, кабель для проводки постоянного тока состоит из двух проводов, в отношении которых не используются такие понятия, как нулевая и фазная жила. В конструкцию кабеля входят лишь две шины с противоположным зарядом, которые иногда называют просто «плюсом» и «минусом».
Принятая маркировка проводов требует, чтобы плюсовой полюс в такой сети был обозначен красным, а минусовой – синим цветом. Нулевой контакт, обозначаемый на схемах М, окрашивается в голубой цвет.
Когда двухпроводная сеть подключается к трехпроводной, необходимо, чтобы цвета ее проводов или шин точно соответствовали цвету контактов питающей электросети, к которым они подсоединены.
Ручная цветовая разметка
Применяется в тех случаях, когда при монтаже приходится использовать провода с жилами одинаковой расцветки. Также часто это происходит при работе в домах старой постройки, в которых монтаж электропроводки производился задолго до появления стандартов.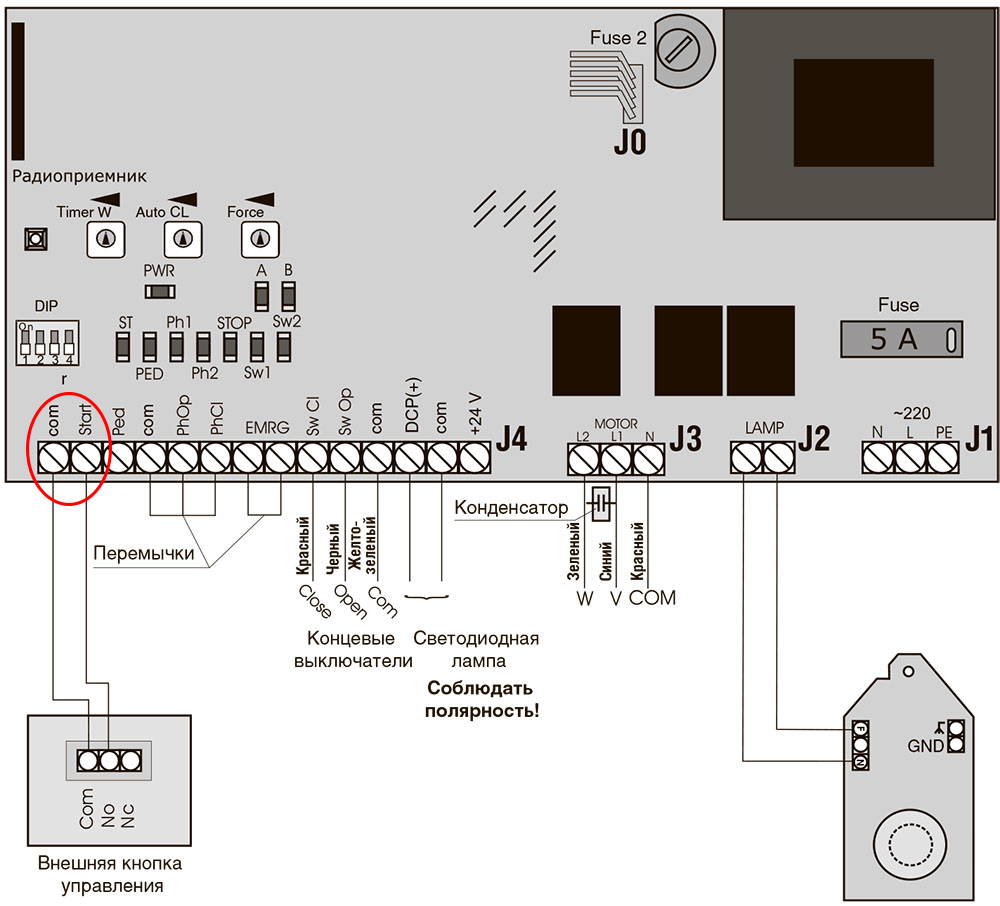
Опытные электрики, чтобы не было путаницы при дальнейшем обслуживании электроцепи использовали наборы, позволяющие промаркировать фазные провода. Это допускается и современными правилами, ведь некоторые кабели изготавливаются без цветобуквенных обозначений. Место использования ручной маркировки регламентировано нормами ПУЭ, ГОСТа и общепринятыми рекомендациями. Она крепится на концы проводника, там, где он соединяется с шиной.
Разметка двужильных проводов
Если кабель уже подключен к сети, то для поиска фазных проводов в электрике используют специальную индикаторную отвертку – в ее корпусе есть светодиод, который светится, когда жало устройства касается фазы.
Правда эффективной она будет только для двухжильных проводов, ведь если фаз несколько, то определить где какая индикатор не сможет. В таком случае придется отключать провода и использовать прозвонку.
Далее понадобится набор специальных трубок с термоусадочным эффектом или ленты для изоляции, чтобы разметить фазу и ноль.
Стандарты не обязывают делать такую разметку на электропроводниках по всей их длине. Допускается отметить её лишь в местах стыков и соединения нужных контактов. Поэтому, при возникновении необходимости нанести метки на электрокабели без обозначений, нужно заранее приобрести материалы, для их разметки вручную.
Число используемых расцветок зависит от применяемой схемы, но главная рекомендация все же есть – желательно использовать цвета, исключающие возможность путаницы. Т.е. не применять для фазных проводов синие, желтые или зеленые метки. В однофазной сети, к примеру, фазу обычно обозначают красным цветом.
Разметка трехжильных проводов
Если надо определить фазу, ноль и заземление в трехжильных проводах, то можно попробовать сделать это мультиметром. Прибор устанавливается на измерение переменного напряжения, а затем щупами аккуратно коснуться фазы (его можно найти и индикаторной отверткой) и последовательно двух оставшихся проводов. Далее следует запомнить показатели и сравнить их между собой – комбинация «фаза-ноль» обычно показывает большее напряжение, нежели «фаза-земля».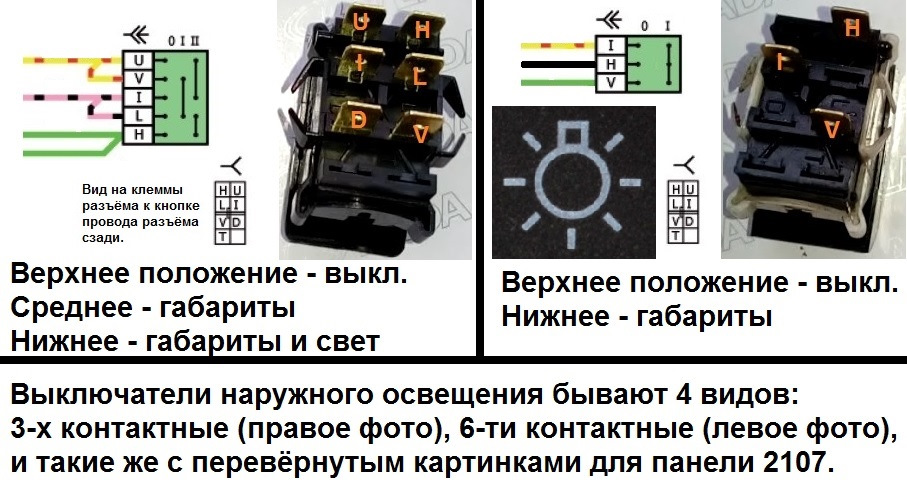
Когда фаза, ноль и земля определены, то можно наносить маркировку. По правилам, для заземления применяется провод цветной желто зеленый, а точнее жила с такой расцветкой, поэтому его маркируют изолентой подходящих цветов. Ноль, отмечается, соответственно, синей изолентой, а фаза любой другой.
Если же при профилактических работах выяснилось, что маркировка устарела, менять кабеля не обязательно. Замене, в соответствии с современными стандартами, подлежит только электрооборудование, вышедшее из строя.
Правильная разметка проводов это обязательное условие качественного монтажа электропроводки при проведении работ любой сложности. Она значительно облегчает как сам монтаж, так и последующее обслуживание электросети. Чтобы электрики «разговаривали на одном языке», созданы обязательные стандарты цветобуквенной маркировки, которые схожи между собой даже в разных странах. В соответствии с ними L – это обозначение фазы, а N – ноля.
Видео по схеме установки двойного выключателя
Для более подробного ознакомления о том, как подключить двухклавишный выключатель, рекомендуем просмотреть видео:
Как видите, выключатель ставиться в разрыв фазного провода, идущего к светильнику. Поэтому в подрозетнике с электропроводкой под одноклавишный выключатель, располагается два провода.
Первый, назовем его «А», идёт к выключателю из электрощита и всегда находится поднапряжением.Второй, назовем его «B», идёт от выключателя к светильнику.
Когда вы нажимаете клавишу выключателя – проводники «А» и «B» соединяются, напряжение беспрепятственно идёт к светильнику и лампы в нем загораются. Соответственно при опускании клавиши, контакт разрывается и свет гаснет.
Теперь, если вспомнить основные обозначения в электрике, которые мы рассматривали ЗДЕСЬ (их не так много, советую ознакомится на будущее), становится понятным, что значит маркировка «L» на контакте выключателя.
Обозначение «L», на выключателе, указывает на контакт для подключения фазного провода. Того самого провода «А» в нашей схеме, который идёт от электрощита и всегда находится под напряжением.
Того самого провода «А» в нашей схеме, который идёт от электрощита и всегда находится под напряжением.
Определить, какой из проводов в подрозетнике необходимо поместить в клемму L выключателя света довольно просто — достаточно проверить, например, индикаторной отверткой, на каком из проводников есть напряжение – тот и будет искомым фазным проводом «А».
В оставшийся, свободный, контакт одноклавишного выключателя, который может быть маркирован по-разному: L1, L`, стрелочкой, «1» или вообще никак, подключается провод «B» из нашей схемы, который идёт непосредственно к выключателю.
Довольно подробно о том, как правильно подключить одноклавишный выключатель, с описанием не только его контактов и порядка соединения проводов, а всего процесса монтажа, вы можете ознакомиться ЗДЕСЬ.
Если же вам при осмотре клемм выключателя света, кроме обозначения L и L1 встретились еще контакты, имеющие какие-то маркировки, то скорее всего вы имеете дело двух- или трех-клавишным выключателем.
При определении назначения контактов, например, двухклавишного выключателя работает та же логика, давайте рассмотрим его схему.
При подключении двухклавишного выключателя используется три провода, которые доступны при монтаже в подрозетнике, это:
«А» — фазный провод, идущий от электрощита и находящийся всегда под напряжением. Подключается к контакту L двухклавишного выключателя.
«B» — проводник,идущий к первому светильнику, либо же включающий первый режим работы люстры. Подключается к клемме L1, L` или просто «1» выключателя света.
«C» — провод, идущий ко второму светильнику или включающий второй режим работы той же люстры. Подключается к клемме L2, L« или просто «2» выключателя света.
Думаю, теперь общий принцип маркировки всех выключателей света вам понятен. Подробнее о том, как подключить двухклавишный выключатель, какие и куда провода следует подсоединить, описано ЗДЕСЬ.
Контакт L – это всегда место для подключения фазного провода.
Остальные же контакты (L1, L2, L3), чаще всего пронумерованные по порядку, относятся к соответствующим клавишам выключателя, нажатие которых зажжёт светильник, подключенный к клемме этой клавиши.
Определить, какой из проводов отвечает за включение какого из светильников, без специального оборудования, довольно сложно. Поэтому обычно их связь выявляется экспериментально.
Поочередно соединяя свободные проводники с фазным проводом в подрозетнике, вы сможете заметить какие светильники зажигаются. Другими словами, вы можете подключить выключатель проихвольно (кроме клеммы «L») и, если клавиши перепутаны, просто переставить местами провода в клеммах L2 и L3, если выключатель двухклавишный.
Если же контактов для подключения три или четыре, а выключатель света одноклавишный, или же контактов шесть, а выключатель двухклавишный, то тогда, вы скорее всего держите в руках один из видов переключателей.
Схему подключения проходного переключателя — три контакта для подключения проводов у одноклавишного устройства вы можете посмотреть ЗДЕСЬ. Двухклавишного переключателя — шесть клемм для подключения проводов ТУТ.
Двухклавишного переключателя — шесть клемм для подключения проводов ТУТ.
Схему подключения перекрестного переключателя – четыре контакта для подключения проводов у одноклавишной модели – ЗДЕСЬ.
Как определить L, N и PE?
Итак, представим такую ситуацию: в процессе ремонта бытовой электрической сети случилось так, что каждый из проводников обладает одним и тем же цветом. Как же определить, какой провод L, какой N, а какой PE?
Если однофазная сеть имеет всего 2 жилы, то проблему можно разрешить специальной индикаторной отверткой. С её помощью проще простого определить, где именно ноль, а где фаза. О том, как использовать индикаторную отвертку мы уже рассказывали. Для начала необходимо отключить подачу электричества на щитке.
После нужно зачистить два проводника и развести их в противоположные стороны. Теперь можно включить подачу электричества и посредством индикатора определить L и N. Если при контакте с жилой загорится лампочка, значит это фаза, в то время как не загоревшаяся лампочка будет подразумевать ноль.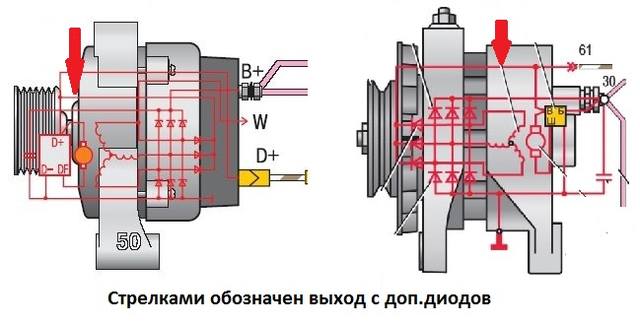
В случае, если в электропроводке присутствует заземляющий провод, нужно будет прибегнуть к такому электроизмерительному оборудованию, как мультиметр. Он оснащается двумя щупальцами. Для начала необходимо установить показатель измерения переменного тока на показатель более 220 Вольт. Одна из щупалец фиксируется на фазном контакте. При помощи другого щупальца определяется заземление и ноль.
При соприкосновении с нулем на электроизмерительном приборе отобразится значение напряжения в районе 220 Вольт. Если же вы прикоснетесь к «земле» – данный показатель будет ниже. Более детальная инструкция по эксплуатации данного прибора была предоставлена в отдельной статье, с которой мы советуем ознакомиться!
Существует альтернативный метод определения. В случае отсутствия индикаторной отвертки и мультиметра под рукой, можно постараться вычислить цвет проводов по их изоляции
В этом случае важно помнить, что синяя оболочка в любом случае всегда будет нулём. В случае даже самой нестандартной маркировки, окрас нуля не меняется
В случае определения остальных двух жил всё будет несколько сложнее.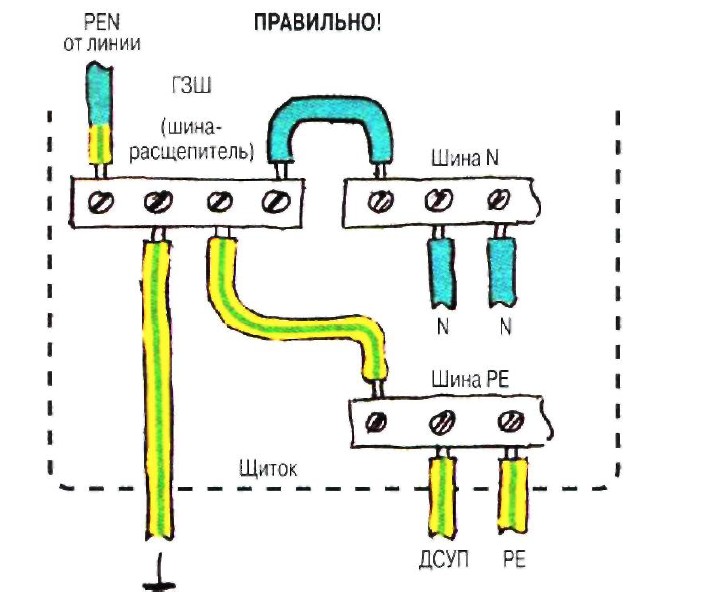
В первом возможном варианте вы видите оставшийся цветной, а также черный или белый контакт. Раньше землю обозначали изоляцией черного, либо белого цвета. Вполне вероятно, что это именно она, а оставшийся цветной – фаза (L).
Во втором возможном варианте также откидываем ноль, концентрируясь на красном и черном (или белом) проводе. Если изоляция имеет белый цвет, то по ПУЭ – это фаза. Тогда, оставшийся красный является землей.
Обратите внимание на то, что последний метод крайне опасен. Если Вы решили прибегнуть к нему, обязательно сделайте для себя пометки, чтобы в процессе ремонта розетки либо люстры не получить электрический удар!. В цепи постоянного тока, маркировка представлена черным (минус), а также красным (плюс) окрасом изоляции
В случае трехфазной сети для каждой фазы имеется свой индивидуальный окрас: фаза A — желтая, B — зеленая, а C — красная. Ноль также синим, а заземление будет иметь желто-зеленый окрас
В цепи постоянного тока, маркировка представлена черным (минус), а также красным (плюс) окрасом изоляции. В случае трехфазной сети для каждой фазы имеется свой индивидуальный окрас: фаза A — желтая, B — зеленая, а C — красная. Ноль также синим, а заземление будет иметь желто-зеленый окрас.
В случае трехфазной сети для каждой фазы имеется свой индивидуальный окрас: фаза A — желтая, B — зеленая, а C — красная. Ноль также синим, а заземление будет иметь желто-зеленый окрас.
В случае кабеля на 380 В провод A будет белым, B черным, а C красным. Нулевой защитный и рабочий проводники имеют аналогичные с последним случаем маркировки.
Список источников
- guru220v.ru
- remontkvartiri.me
- electricremont.ru
Поделитесь с друзьями!
Клеммы в электрощитах: Зачем они нужны. Подключение нулей .NLP перемычками – CS-CS.Net: Лаборатория Электрошамана
Клеммы для удобного подключения отходящих линий в щите
Всем привет! Так случилось, что несколько раз по почте меня разный народ закидал двумя вопросами: нахрена я ставлю клеммы в щитах, если там можно кабели подключить напрямую к дифавтоматам и как правильно подключать ноль на клеммы . NLP, если пользоваться перемычками.
NLP, если пользоваться перемычками.
Внимание! В Августе 2018 года ABB передала завод EntrElec, который производит клеммы, фиксаторы и кросс-блоки фирме TE Connectivity. Всё в порядке: все артикулы клемм и все клеммы остаются, просто называться они будут вместо «ABB D4/6.NLP» — «TE (EntrElec) D4/6.NLP». Все коды заказа сохраняются. Читайте про это (и следите за новостями) в моём посте со всей информацией.
Так же TE сделали навигатор в Excel по их клеммам с возможностью отбора по параметрам. Файлик я выложил себе на хостинг вот тут: TE-Entrelec-Connectors-Selection.xlsx (~1,3 мб).
В принципе вопросы-то простые и лёгкие, но так вышло, что одному парню по почте я нашёл аж пять идей, почему нужны клеммы в щите и подумал — а чего ж этот ответ пропадает: Ну и решил черкнуть пост. Свежих фоток тут не будет, я натаскаю их из других постов, потому что тут они нам нужны только для иллюстрации некоторых моментов.
И всё же детально разжёвывать некоторые моменты я не буду. Вы же помните, что я заставлю всех думать, подавая нужные идеи, а не выкладываю готовые решения для повторюшек?
Вы же помните, что я заставлю всех думать, подавая нужные идеи, а не выкладываю готовые решения для повторюшек?
Что почитать (все ссылки по клеммам)
Да-да. Соберу все ссылки в кучу, чтобы можно было к ним потом обращаться и те, кто найдёт этот пост, имели всё под рукой:
Если попробовать пояснить кратко, что клеммы на DIN-рейку состоят из вот таких вот штуковин:
- Сами клеммы. На большие сечения они выпускаются отдельно (есть клеммы на 70 и даже 240 квадратов О_о), а в некоторых случаях выпускаются в виде одной клеммы, но с несколькими контактами (L, N, PE или просто три контакта для любых целей).
- Клеммы предназначены для того, чтобы крепить их на DIN-рейку, потому что PE они берут именно с неё (у них нет контаков для ввода PE в клемму). Клеммы небольших сечений нащёлкиваются на рейку, а клеммы больших сечений затягиваются на ней винтом, чтобы иметь хороший контакт и держать ток замыкания на землю.
- Обычно одна из боковых сторон клемм закрыта, а другая — открыта.
 Это делается для того, чтобы клеммы были как можно уже по ширине и не занимали много места. Изоляция клеммы рассчитана и спроектирована на такой способ применения.
Это делается для того, чтобы клеммы были как можно уже по ширине и не занимали много места. Изоляция клеммы рассчитана и спроектирована на такой способ применения. - Для того, чтобы закрыть край клеммной сборки (последнюю клемму, которая будет стоять открытой стороной наружу) используются торцевые изоляторы. Также этими изоляторами можно отделить разные логические группы клемм друг от друга.
- Для клемм выпускаются перемычки. Это специальные пластинки, которые соединяют соседние клеммы между собой. Перемычки для каждого вида клеммы будут своими, потому клеммы разного вида и сечения имеют свою ширину и ширина перемычки тоже должна быть такой же, как и у клеммы.
Перемычки можно ставить только на те клеммы, которые стоят вплотную друг к другу без торцевых изоляторов. Также перемычки обычно рассчитаны на тот же ток, на который расчитана одна клемма (про это мы ещё поговорим). - Клеммные сборки удобно фиксировать специальными фиксаторами.
 Они обычно имеют большую высоту для того, чтобы прижимать клемму не снизу у рейки, а по всей высоте. У ABB это фиксаторы BAM4. А ещё у них есть специальные вставочки PEAD, в которые можно вложить бумажку с подписью ряда клемм.
Они обычно имеют большую высоту для того, чтобы прижимать клемму не снизу у рейки, а по всей высоте. У ABB это фиксаторы BAM4. А ещё у них есть специальные вставочки PEAD, в которые можно вложить бумажку с подписью ряда клемм.
1. Где удобно применять клеммы в щитах и чем они могут быть полезны?
Мой путь с клеммами был тернист и долог. Сначала я не знал, нафига их применять, а потом постепенно стал делать на них небольшие вставки для автоматики в щитах. Например, если у нас есть какой-то кабель управления, то вместо того чтобы объяснять заказчику: «…первый провод надо подсунуть вместе с фазой от автомата Q12, второй — на контакт А1 реле K15, а третий — на вот эту нулевую шинку» можно просто поставить клеммы для этого кабеля, пронумеровать их, а в щите соединить всё гибким проводом. И пусть заказчик подключается к клеммам.
Постепенно эта идея росла и крепла и когда я делал свой первый огромный рэковый щиток, то все линии автоматики я там сделал на клеммах. Вот так это выглядело:
Клеммные блоки для подключения автоматики освещения
Но силовые линии у меня там шли без клемм. Для них я предполагал использовать шинки N/PE. Вот они тут видны снизу фотки:
Для них я предполагал использовать шинки N/PE. Вот они тут видны снизу фотки:
Вид на собранную часть автоматики щитка (импульсные реле)
В результате подключение щита выглядело вот так:
Подключение силовой части: не помешали бы и клеммники
Спрашивается… если пошли такие огромные щиты (по тем меркам), то почему не попробовать заказать на щит специальные клеммы, которые значились в каталоге как «…для электрощитов»? Это были клеммы D4/6.NLP, к которым сразу целиком удобно подключается кабель сечением до 3х2,5.
И вот как выглядел на них мой первый щит на таких клеммах:
Почти все подключения закончены!
Кабелей было больше, потому что я вывел сюда все линии (и силовые тоже), а клеммы заняли места меньше. И вот после этого у меня и свершился скачок! Я на опыте понял, что клеммы упрощают работу со щитами, если щиты большие или сложные по автоматике. А потом ко мне подтянулся первый заказчик, который попросил сделать щит с клеммами для того, чтобы тупые рабочие ничего в щите не трогали, кроме этих клемм. И — закрутилось!
И — закрутилось!
Итак, давайте подбивать мысли о том, где могут сгодиться клеммы.
Во-первых, клеммы удобно ставить, если надо подключить к щиту толстые бронированные кабели или короткий кабель ввода. Вместо того, чтобы тащить кабель, к примеру, 4х16, к вводному рубильнику щита (который находится вверху), можно поставить клеммы снизу щита, а дальше повести всё мягкими проводами. Вот так было сделано в щите в Поварово:
Клеммы ввода сети, генератора, стабилизаторов. Тоже подписаны
А вот так сделано в щите в СИТИ для кабелей 4х70 и 5х25:
Клеммы для подключения силовых кабелей на 70 кв.мм.
Во-вторых, если щит сам по себе получается большим (шкаф типа B/TwinLine), то клеммы удобно ставить для того, чтобы… экономить КАБЕЛЬ! Потому что обычно мы ставим клеммы с той стороны шкафа, с какой в него входят кабели. В этом случае запас кабелей нам будет нужен небольшой — только до клемм (например, 0,5 метра). А если вести кабели напрямую до автоматов, то запас нужен будет минимум полторы высоты шкафа (например, 2 метра).
Подсчитаем? Положим, приходит к нам 90 кабелей. Если сделать всё на клеммах, то наша разница запаса будет 2-0,5 = 1,5 метра на каждый кабель! 90 х 1,5 = 135 метров! Да это ж целая бухта кабеля, блин! А ведь клеммы дают и просто очень удобное подключение, если ещё и позаботиться о свободном месте около них:
Все отходящие линии щита сделаны на клеммах для удобства
В-третьих, про автоматику уже сказали. Как только в щите заводятся линии для каких-нибудь кранов, кнопок, ламп света — то без клемм не обойтись.
В-четвёртых, иногда клеммы нужны из-за конструкции самого щита. Например, когда у меня заказывают щиты на голой WR-раме, то там не будет места, по которому провода от кабелей можно дотащить до автоматов — только за DIN-рейками пропихивать.
Поэтому в таких щитах я всегда ставлю клеммы. Вот красивая фотка того, как Кирич подключал мой щит на WR-раме:
Провода ради прикола сложены петлями и выровнены по лазерному уровню
В-пятых, клеммы нужно использовать, когда нам надо объединить несколько кабелей в один.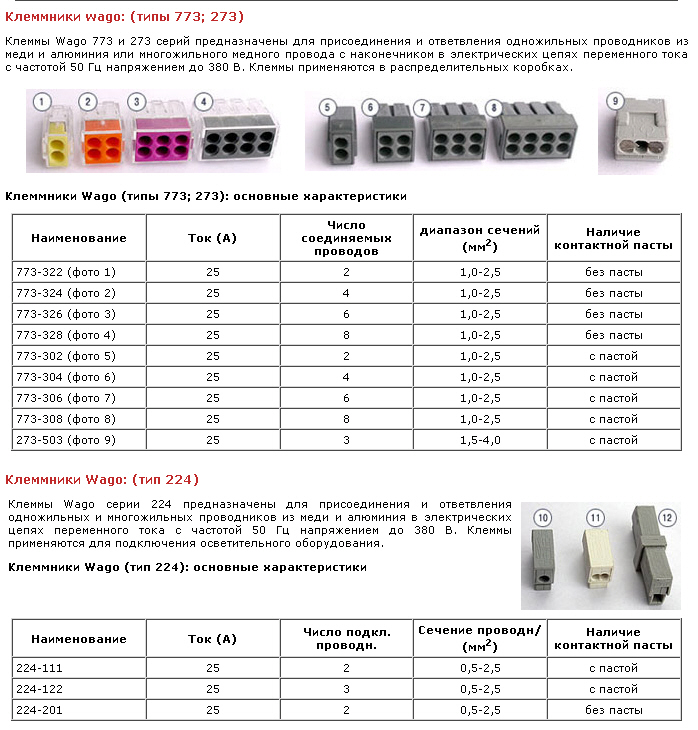 Некоторые любят делать проводку без распаечных коробок, но не монтажом в подрозетниках, а тем, что сводят кабели от каждого блока розеток в щит. Тогда может получиться, что из какой-нибудь комнаты идёт пять кабелей, которые должны оказаться на одном автомате.
Некоторые любят делать проводку без распаечных коробок, но не монтажом в подрозетниках, а тем, что сводят кабели от каждого блока розеток в щит. Тогда может получиться, что из какой-нибудь комнаты идёт пять кабелей, которые должны оказаться на одном автомате.
Тут на помощь и придут клеммы с перемычками. Вот так это выглядит в щите на Ополченцы:
Клеммы для подключения отходящих линий
Видите, к некоторым клеммам провода не подходят? Вот там и стоят перемычки! А раз уж мы начали делать что-то на клеммах — то имеет смысл совместить все технологии клемм в одну кучу и сделать весь щит на клеммах целиком, вот так:
Пространство внутри корпуса для кабелея около клемм
Ещё и свободное место осталось для разделки кабелей (так и должно быть).
Ну и в-шестых, да — тупые рабочие. Иногда меня заказчики так прямо и просят: «У меня рабочие тупые! Можно мне всё сделать на клеммах, чтобы они никуда, кроме клемм, не лазили?».
2.
 Клеммы надо применять с умом. Какие косяки с ними бывают?
Клеммы надо применять с умом. Какие косяки с ними бывают?Я бы выделил два косяка, которые надо учитывать, если вы хотите сделать щит на клеммах.
Косяк первый — не забывайте считать ширину клемм и проверять, влезут ли они на DIN-рейку. А ещё не забывайте учитывать ширину фиксатора BAM4, если вам надо будет разделить клеммные сборки между собой для красоты.
CS CRM: Калькулятор ширины клеммы на DIN-рейку
Для себя я накрутил 1Ску так, чтобы она умела считать ширину нужных мне клемм с запасом и завёл такое негласное правило: если клеммы еле-еле вместились на рейку в 1Ске, то в реале они гарантированно влезут в щит.
А второй косяк — не совсем косяк, а особенность. Всё-таки рано или поздно вы поймёте, что около клемм надо оставлять место для кабелей. Вот пример плохого щита на СтароВолынскую:
Плохо сделано: клеммы слишком близко к краю щита
Здесь заказчик настоял на том, что надо использовать щиты простой серии AT/U, и они получились забитыми под завязку. В итоге для клемм места почти не осталось (а там автоматика была — клеммы были нужны), и расстояние от края клемм до корпуса щита было всего лишь около 2-3 сантиметров.
В итоге для клемм места почти не осталось (а там автоматика была — клеммы были нужны), и расстояние от края клемм до корпуса щита было всего лишь около 2-3 сантиметров.
Вот как это выглядит:
Расстояние от клемм до края корпуса
Кайф использования клемм как раз в том, чтобы оставить около них свободное место. Тогда в этом месте можно устроить любое (и некрасивое) месиво для кабелей! То есть, тогда щит делится на две части: чистенький и аккуратный щит, и место для кабелей, где можно делать что хочешь и как хочешь.
Вот как должно быть:
Пространство для кабелей внутри щита
Для себя я взял такое правило: если клеммы занимают одну DIN-рейку — то ещё одну DIN-рейку мы оставляем как свободное место. То есть, берём модуль для клемм двойной высоты и одну рейку просто не ставим. Ну а если клеммы будут занимать две рейки — то (если шкаф позволит) берём модуль тройной высоты.
На этом, наверное, про особенности монтажа клемм — всё.
3.
 Фишка с нулями от УЗОшек на клеммах .NLP.
Фишка с нулями от УЗОшек на клеммах .NLP.И теперь я снова попробую разъяснить фишку, которую уже пояснял в посте про перемычки для клемм. Речь идёт о том, что когда мы узнаём о том, что для клемм есть перемычки, то нам приходит в голову отличная мысль: «Во! Тогда раз у меня под УЗО стоит десять автоматов — то я ща на клеммы поставлю перемычку и подкину ноль одним проводом»!
Дальше обычно мыслят так:
— Это… так… десять автоматов. Каждый по 16А. Это ж какое сечение нуля должно быть? Ну.. не меньше вводного кабеля. На десять квадратов.
— Ой.. а как десять квадратов запихнуть в клемму на 2,5..4 квадрата? Чё оно не лезет-то?
И дальше обычно пишут мне или где попало на блоге или на мыло =) Правильно! Если «оно» не лезет — то так и задумано для того, чтобы заставить вас задуматься. И первый вопрос, который надо себе задать, будет такой: «А с чего ты вообще решил, что эта перемычка должна выдержать адский общий ток нулей?»
Всё верно! Она его и не должна держать! Обычно перемычка рассичтана на тот же ток, что и одна клемма. И поэтому перемычки можно применять тогда, когда их ток ограничен чем-то извне (автоматом) и не будет больше тока одной клеммы (для .NLP это около 32-38А; берём худший вариант на 32А).
И поэтому перемычки можно применять тогда, когда их ток ограничен чем-то извне (автоматом) и не будет больше тока одной клеммы (для .NLP это около 32-38А; берём худший вариант на 32А).
Распишем разные случаи:
- Если клеммы используются для того, чтобы размножить линию с одного автомата на несколько, то все эти несколько клемм будут стоять под одинм автоматом на 10..16А — поэтому перемычку можно применять без проблем (и нужно).
- Если клеммы используются для того, чтобы управлять светом или чем-то ещё. Например, фаза с автомата пошла через три импульсных реле на три клеммы. Тогда через эти три клеммы общая нагрузка никогда не будет больше номинала автомата. И значит перемычку можно применять.
- Если суммарная сумма номиналов автоматов, которые выведены на клеммы и чьи нули вы хотите соединить, больше номинала перемычки — то всё, перемычку просто так нельзя применять. Но именно в этом случае вам понадобится провод большого сечения, вы его не запихаете и задумаетесь =)
Последний вариант я решаю хитро. Я СЧИТАЮ, сколько автоматов у меня окажется под общим нулём и перемычку на нули всё равно ставлю, но протягиваю этот ноль несколькими отдельными проводами. Вот примеры таких расчётов:
Я СЧИТАЮ, сколько автоматов у меня окажется под общим нулём и перемычку на нули всё равно ставлю, но протягиваю этот ноль несколькими отдельными проводами. Вот примеры таких расчётов:
- УЗО на 40А, стоит 4 автомата по 10А (свет). Значит всего будет 40А. Ставим перемычку, и нули тащим тремя проводами по 2,5 квадрата. Одна из клемм питается за счёт соседней перемычки.
- УЗО на 40А, 4 автомата по 16А (розетки). Ток всё равно ограничен 40А (раз узо выбрано на 40 по вводному автомату). Но для надёжности в этом случае я протащу четыре нуля. Перемычка тут ставится для того, чтобы уравнять токи по параллельным проводам.
А вот если под одним УЗО стоит дохрена автоматов (например, 10) по 16А — то тут может быть целая куча вариантов. Считаем, что УЗО у нас на 63А для однофазных щитов:
- Я бы поставил блок BRU/DBL (или другой) и всё равно умудрился протащить все 10 нулей отдельными проводами. Линии силовые — нехрен экономить!
- Можно перейти на провода по 4 квадрата и запихать их в клеммы через один, потому что всё равно УЗО выбрано на 63А и суммарное сечение может быть не больше 10 квадратов внутри щита.
- Можно разбить эти линии на несколько УЗО (это будет лучший вариант из всех)
Как запихать несколько проводов в один зажим УЗО? Мне понравилось использовать наконечники НШВИ(2). Оказалось, что в такие наконечники влезает некоторое количество проводов по 2,5 квадрата. Например в НШВИ(2) на 10 квадратов можно запихать 5-6 проводов по 2,5. В НШВИ(2) на 6 квдаратов — 4 провода по 2,5.
Вот так это выглядит в щитах. Из УЗО выходит пачка проводов нулей:
Монтаж щита внутри: с УЗО и автоматов — на клеммы
А потом приходит на клеммы подряд:
Клеммы D4/6.NLP для подключения отходящих линий (сверху)
Когда я собираю щит, я прикидываю примерную длину одного провода, нарезаю их пачку и опрессовываю в один НШВИ. А потом по месту подрезаю их по нужной длине у каждой клеммы и закручиваю туда. Вот и все секреты!
Схема подключения датчика движения, управление светом
Схемы подключения датчика движения (правильнее его называть датчиком управления освещением или светом) на сайтах соответствующей тематики не публикует только ленивый, поэтому написать что — то оригинальное на эту тему достаточно тяжело.
Поэтому здесь я попробую акцентировать Ваше внимание на некоторых моментах, которые позволят подключить датчик движения осознанно, что бывает необходимо при возникновении некоторых нестандартных ситуаций.
Для начала предлагаю структурную схему датчика движения (рисунок 1).
Здесь:
- BL — собственно сам датчик движения,
- S — контакт управления освещением,
- N — «нулевой» провод осветительной сети,
- L — «фаза»,
- A — клемма подключения осветительных приборов.
Датчик движения действует по принципу обнаружения изменений интенсивности ИК (инфракрасного излучения) в зоне его действия, то есть обнаруживает движущийся объект, который пересекает специальным образом сформированные сектора зоны обнаружения.
То есть, если Вы будете пребывать в зоне обнаружения датчика без движения или интенсивность движения будет недостаточной он посчитает, что Вас там нет. Конечно, сначала, когда кто то входит в зону действия включение происходит, но стоит ему приостановить движение, датчик (соответственно свет) выключится.
Для предотвращения подобных явлений в датчике должна быть предусмотрена регулировка задержки времени отключения: TIME.
Далее, ИК датчик имеет разную чувствительность при движении объекта обнаружения в разных направлениях, кроме того он может реагировать на конвенционные потоки (от нагревательных приборов), различного рода засветки, например фарами автомобилей (что следует учитывать при его установке).
Поэтому необходим орган регулировки чувствительности SENS.
Поскольку рассматриваемые датчики предназначены для управления светом то включаться они должны при недостаточной освещенности. Для этого существует регулировка LUX.
Для того, чтобы правильно выбрать датчик движения следует знать такие его характеристики как:
- угол обзора — для вариантов настенной установки он может достигать 1800, потолочной — 3600,
- дальность действия,
- максимальная мощность или ток, коммутируемые управляющим контактом,
- варианты исполнения (настенный, потолочный, внутренней или наружной установки).
ВАРИАНТЫ ПОДКЛЮЧЕНИЯ
- это стандартная схема, комментарии, думаю, излишни,
- в этой схеме присутствует обычный электрический выключатель (S), который позволяет включать освещение независимо от состояния датчика,
- при необходимости подключить нагрузку большой мощности можно использовать электромагнитный пускатель или реле (КМ), главное чтобы его катушка была рассчитана на напряжение 220В, а контактная группа (КМ1) соответствовала мощности нагрузки,
Следующая схема подключения может использоваться когда зона действия одного датчика недостаточна для контроля освещаемой территории. Здесь может быть подключено любое количество датчиков.
Кроме того, на примере этой схемы хочу предостеречь от параллельного подключения нескольких датчиков для увеличения коммутируемой контактами мощности. Конечно, два контакта, например, будут управлять вдвое большей нагрузкой, но при условии, что они сработают строго одновременно. На практике такого не будет никогда.
Несколько слов про неправильные подключения и их последствия: 1. Если поменять местами подключение L и N ничего страшного не произойдет, за исключением того, что даже в выключенном состоянии датчика фаза будет присутствовать на осветительном приборе. Это отрицательно скажется на безопасности, но не работоспособности.
2. Если один из сетевых проводов подключить вместо штатного места на клемму А, кроме того, что датчик движения откажется работать (но не перегорит) ничего не произойдет (см.рис.1). Правда, если по причине неисправности датчика контакты управления будут находиться в замкнутом состоянии возможно короткое замыкание. Поэтому перед установкой прозвоните датчик тестером.
3. Если подключить неправильно лампу она либо не будет гореть (клемма А и L), либо будет гореть постоянно (клеммы L и N), но второй вариант сразу будет заметен по причине незадействованной клеммы А.
© 2012-2020 г. Все права защищены.
Представленные на сайте материалы имеют информационный характер и не могут быть использованы в качестве руководящих и нормативных документов
Как подключить термостат к газовому котлу
Ручная и автоматическая регулировка температуры в помещении от современных источников отопления производится при помощи терморегуляторов или термостатов.
Они бывают механическими и электронными. У каждого из них есть свои плюсы и минусы.
- термостат для водяного отопления
- для электрического теплого пола
- для газовых котлов или бойлеров
По своему внешнему виду они все очень похожи друг на друга, однако схемотехника у них отличается.
Давайте подробнее рассмотрим особенности подключения терморегулятора к газовому котлу, его схему, ошибки и возможные проблемы при эксплуатации подобных устройств.
Преимущества использования терморегулятора
Чтобы правильно выбрать нужную модель термостата переверните его и внимательно изучите обратную сторону. А именно — схему, надписи и контактные колодки.
Если там будет указано “Dry Contact” (сухой контакт) – вы сделали правильный выбор. Так обозначаются терморегуляторы, предназначенные для работы с газовыми котлами.
Термин “сухой контакт” говорит, что в любом состоянии напряжение на этих зажимах отсутствует. То есть, подача питания 220V на колодки L и N термостата, никак не будет связана с сухими контактами, и напряжение туда никогда не попадет.
В отличие от терморегуляторов тёплого пола, как на схеме ниже. У них контакты подачи питания (3-4) и нагрузки (1-2) соединены напрямую между собой.
Зачем вообще нужна эта штука для газового котла? Без такого программатора, котел после выхода на заданную температуру будет работать с задержкой примерно в 3 минуты.
Включился, поработал, набрал температуру и потух. Через три минуты заново включается и все повторяется по новому циклу. За сутки устройство будет запускаться около 200-240 раз.
Таким образом, за весь отопительный сезон газовый котел включится-отключится 40 000 раз. Подключив терморегулятор, вы в несколько раз сокращаете эти коммутации, добиваясь тем самым не только экономии газа, но и уменьшаете износ отопительного оборудования.
Какую температуру выставлять?
Логика работы здесь следующая. В заводских настройках котел греет воду по температуре теплоносителя.
Устанавливая выносной термостат, мы тем самым даем ему команду, греть воду не так как хочется котлу, а согласно настройкам терморегулятора, т.е. на определенную температуру в том или ином помещении.
При нормальном утеплении дома и минимальных теплопотерях, газовый котел с терморегулятором в сутки будет работать всего 3-4 часа.
Если термостат после установки никак не повлиял на время работы котла, то скорее всего на газовом приборе выставлена меньшая температура, чем необходимо. Датчик регулятора просто не успевает прогреться до нужного значения и сработать, в то время как t теплоносителя уже достигла заданного порога.
В инструкциях отдельно прописывают минимальную t на котле при использовании внешнего термостата. Как правило, она должна быть не меньше 65 градусов.
Изначально на отопительном приборе рекомендуют выставлять расчетную температуру, которая полностью покрывает теплопотери здания. Когда эти теплопотери не известны, для стандартной системы отопления берут значения от 60 до 70С.
Если вы проживаете в относительно теплом климате, и зимой температуру в батареях не поднимаете выше 45С, для работы с термостатом ее все равно придется увеличить.
Некоторые задаются вопросом, а какой тогда смысл в установке регулятора и как это приводит к экономии?
Из-за того, что температура получится больше, ничего страшного нет:
- во-первых, котел меньше тактуя, быстрее прогревает систему
- во-вторых, при большей температуре теплоносителя тепло в комнатах держится дольше
- да и максимальное КПД батарей наблюдается именно при t 65С-70С, а не при +45С
Куда и где подключать провода на газовом котле?
Для подключения термостата от клеммной колодки котла к контактам Dry Contact вам нужно подвести два проводника.
По инструкции рекомендуемое сечение медных жил – не менее 0,75мм2.
Длина кабеля до места установки термостата не должна превышать 50 метров.
Необходимые клеммы на самом котле ищите в инструкции завода изготовителя, либо ориентируйтесь по надписям на колодках.
Данные клеммы чаще всего расположены с обратной стороны платы управления.
Однако иногда производители их выносят за пределы платы.
Еще некоторые монтажники при установке подключают сюда кабель и выводят его наружу, чтобы потребитель в дальнейшем не лез к плате и чего-нибудь там не напутал. Так как термостат еще не установлен, концы кабеля закорачивают.
Кстати, с этой выносной колодкой будьте внимательны. Некоторые потребители сталкивались с проблемой заводского перепутывания проводов, как например у Protherm (Протерм).
В итоге все начинало работать как надо, только при обратном подключении.
Изначально между клеммами стоит перемычка. Если перемычку разорвать, то газовый котел просто-напросто перестанет работать.
При подключении термостата заводскую перемычку демонтируют и на ее место заводят кабель, проложенный до места установки терморегулятора. Самое главное никаких внутренних настроек на котле изменять не нужно.
Через эти контакты подается управляющий импульс низкого напряжения (обычно 24В).
У нас есть управляющее устройство (термостат) и ведомое (котел). Между ними по слаботочной линии постоянно происходит передача набора запросов-ответов.
Принцип работы здесь простой – замыкая сухой контакт, мы тем самым разрешаем работу котла, размыкая – прерываем передачу данных и запрещаем. Замыкание и размыкание происходит через реле термостата, в зависимости от выставленной температуры на устройстве и сравнения текущих показателей в комнате.
При срабатывании сначала отключается горелка, затем через определенный промежуток (время выбега насоса) и циркуляционный насос. Хотя в разных моделях алгоритм работы насоса может отличаться.
Один из вариантов алгоритма
Не желательно, чтобы насос отключался сразу же после срабатывания термостата. В этом случае выставьте в меню выбег от 5 до 10 минут. Тогда вся система отопления будет остывать равномерно и промежутки времени срабатывания автоматики увеличатся.
На дорогих моделях, поддерживающих протокол OpenTherm, через выносной программатор можно управлять многими встроенными параметрами котла, а не только его включением-отключением.
Вот наглядные примеры, куда подключать термостат для разных моделей котлов:
BaxiBoschElectroluxVaillantJunkersBuderusViessmanFerroliAtronАльфа КалорImmergas
А что будет, если терморегулятор выйдет из строя? Как в этом случае поведет себя котел?
Все зависит от того, в каком положении останется сухой контакт. Если замкнут — ничего страшного не случится. Котел просто начнет работать по своей последней заданной программе.
Если разомкнут, то отопление не запустится и работать не будет.
Обратите внимание, что на работу горелки в режиме горячего водоснабжения термостат не влияет.
На контакты питания терморегулятора заводится напряжение 220V – фаза и ноль (L и N). Это можно сделать как от отдельной розетки или распредкоробки, так и взять их непосредственно с самого котла.
На регуляторе с сухими контактами без разницы, куда заводить фазу или ноль, а вот на моделях для эл.теплых полов это играет существенную роль в вопросе безопасности эксплуатации.
Так как при неправильном подключении разрываться будет только нулевой проводник, а фаза будет постоянно дежурить на кабеле обогрева.
Механический или электронный термостат
Кстати, для газового котла можно использовать и другой простейший тип регулятора, на который даже не придется подавать напряжение 220В. Например, механический термостат Termec Emmeti или другие подобные модели.
Вот «обычная» схема подключения Termec.
Вам же нужно задействовать только нормально замкнутые контакты 1 и 3, полностью исключив переменку 220В (L и N).
Встроенный датчик при изменении температуры в комнате будет замыкать и размыкать внутренний контакт. Никакого питания ему не нужно. При этом вся логика работы отопления аналогична рассмотренной ранее.
Только не забывайте, что практически у всех механических моделей очень большой гистерезис. Комфортную температуру в помещениях с их помощью не создашь.
Поэтому по возможности выбирайте электронные девайсы с подключением по WiFi. Благо в наше время у китайцев можно найти очень достойные и недорогие варианты.
Например, такие как этот (тысячи довольных покупателей и положительных отзывов). Подробнее
В некоторых моделях есть контакты, обозначенные как NO (нормально открытые), NC (нормально замкнутые) и COM (общий). Кто-то советует подключать через них, а именно через NC и COM.
Однако будьте осторожны, термостат термостату рознь и всегда изучайте инструкцию. Через них может также подаваться переменное напряжения 220V, и вы тем самым запустите фазу на плату управление куда не нужно.
Вот яркий пример таких многофункциональных регуляторов Fluoreon и Beok.
На многофункциональных устройствах, температура в комнате также определяется при помощи встроенного температурного датчика.
Однако у них на корпусе есть клеммы для подключения и внешнего (Sensor). Он чаще всего используется для теплых полов.
Место установки
Как известно, температура воздуха в помещении с традиционными системами обогрева на радиаторах, прогревается неравномерно. Возле пола она ниже, под потолком выше.
Исходя из наличия в терморегуляторах именно встроенного температурного датчика, их высота установки регламентируется.
Размещать такие термостаты нужно на высоте 1,2-1,5м от уровня пола и как можно дальше от источников обогрева, в том числе защищать от попадания прямых солнечных лучей.
Также не рекомендуется ставить термостаты в коридоре или на кухне.
Подключение термостата теплого пола на газовый котел
Зачастую потребители задаются вопросом, а можно ли использовать стандартный терморегулятор от теплых полов на 220В для работы с газовым котлом?
Вдруг вы не нашли специальный термостат с сухими контактами, а модель от теплых полов у вас уже есть в наличии. Такое подключение возможно.
Однако для этого вам понадобится “развязать” напряжение 220V. В большинстве термостатов теплых полов оно приходит на клеммы питания и далее снимается с клемм, отходящих на кабель обогрева. А как говорилось выше, при работе с газовым котлом используются “сухие контакты”, по которым передаются импульсы низкого напряжения.
Если вы напрямую подключите сюда кабель с платы управления котла, то спровоцируете короткое замыкание и выведете из строя свой котел.
Для безопасного подключения в схему придется добавить промежуточное реле или дополнительный контактор с нормально открытыми контактами.
С клемм термостата вместо кабеля теплого пола подаете напряжение 220В на катушку реле (контактора), а уже через его нормально открытые контакты (1-2 или 3-4), подключаете провода на газовый котел.
Принцип работы здесь такой же, как был рассмотрен ранее. Однако имейте в виду следующую особенность.
Так как это термостат все-таки для теплых полов, то скорее всего он будет работать по схеме: датчик воздуха + датчик пола. А значит потребует от вас установки и выставления ограничения температуры по датчику пола, которого у вас не будет.
Поэтому в настройках придется отвязать датчик пола и оставить в работе только датчик воздуха. Как это сделать, подробно читайте в инструкции к своей модели. У разных девайсов могут быть отличия.
Например, у Devi Touch для этого приходится ломать на задней стороне специальную перемычку.
Дабы не иметь всех этих сложностей и проблем, лучше все-таки изначально выбирать правильную модель терморегулятора.
Терморегулятор для коллектора водяных полов
При наличии в отдельных комнатах водяного теплого пола, который может быть запитан от того же газового котла, нужен несколько иной тип терморегулятора. Его необходимо подключать к сервоприводам на распределительном коллекторе.
Обратная сторона такого устройства выглядит следующим образом.
Если у вас сервопривод нормально закрытый, то жилы кабеля заводятся на клеммы 1 (Close или NC) и 3 (N-ноль или COM). Если нормально открытый – 2 (Open или NO) и 3 (N-ноль или COM).
Принцип работы здесь следующий. Когда в системе установлен нормально открытый сервопривод и при этом на питающем кабеле происходит прерывание подачи напряжения, сервопривод открывает проток. В нормально закрытом, ситуация противоположная.
При этом не путайте, это не какое-то реле или эл.магнитный привод, который при подаче напряжения 220В перещелкивается и моментально переходит в другое состояние. Это термоэлектрический привод, который открывает-закрывает клапан постепенно.
Поэтому, если вы захотите испытать работоспособность подобного устройства, не удивляйтесь, что он сразу же не среагирует на подачу сигнала и моментально не перекроет поток.
Подключение питания термостата осуществляется на колодках L и N. На боковых клеммах заводится кабель выносного температурного датчика в полу.
Статьи по теме
Ключ для идентификации клемм генератора
Этот ключ может быть полезен для идентификации клемм на ваших генераторах, поскольку вы можете видеть, что одна и та же функция может иметь разные обозначения в зависимости от марки и модели генератора| Терминал | Определение | Банкноты |
| А | Аккумулятор | Плюсовой провод аккумуляторной батареи |
| B | Аккумулятор | Плюсовой провод аккумуляторной батареи |
| Б + | Аккумулятор | Плюсовой провод аккумуляторной батареи |
| С | Центр | Центральная точка статора Y |
| COM | Компьютер | Подключение к компьютеру |
| Д + | Сигнальная лампа | Происходит от старого «Dynamo Positive» на генератор |
| DFM | Динамо-модуляция поля | Сигнал рабочего цикла от ЭБУ |
| E | Сигнальная лампа | Произошло от слова «возбуждение» |
| F | Поле | Для генераторов с внешней регулировкой |
| FR | Регламент поля | Сигнал напряжения от ЭБУ |
| G | Земля | Отрицательное соединение шасси |
| I | Инидкатор | Контрольная лампа |
| л | Лампа | Контрольная лампа |
| M | Поле | Обнаружен на более старых устройствах с внешним регулированием |
| N | нейтральный | Центральная точка статора, соединенного звездочкой |
| -п. | Импульс | Подключение статора для тахометра |
| R | Реле | Выход для реле заряда и т. Д. |
| S | Sense | Провод датчика напряжения аккумулятора |
| S (FORD) | Статор | Импульс статора.Используется для тахометров |
| т | Тахометр | Импульс статора. Используется для тахометров |
| Вт | Форма волны | Импульс статора. Используется для тахометры |
| Символ | Описание | Символ | Описание | |
|---|---|---|---|---|
| Электролиния / кабель / провод Жила однониточная + Инфо | Линия электропередачи / Линия электропередачи / кабель / Провод Линия электропередачи + Инфо | |||
| Электролиния / кабель / провод Электропроводка однониточная + Инфо | Место соединения в проводнике | |||
| Линии пешеходного перехода без подключения, перекрестный провод Нет контакта / нет соединения | Линии пешеходного перехода без подключения, перекрестный провод Нет контакта / нет соединения | |||
| Бесконтактный переход, перекрестный провод Нет контакта / нет соединения | Соединительные провода | |||
| Соединительные провода | Соединительные провода | |||
| Соединительные провода | Соединительные провода | |||
| Двухпроводная линия | Трехпроводная линия | |||
| Трехпроводная линия | Провод Звездочкой указано количество проводов | |||
| Нейтральный провод + Инфо | Защитный провод + Инфо | |||
| Кабель с защитной линией и нейтралью | Трехфазный, защитный и нейтральный проводник | |||
| Экранированный кабель / Экранированный провод + Информация | Экранированный кабель / Экранированный провод | |||
| Линия направления | Коаксиальный кабель + Инфо | |||
| Линия задержки + Инфо | Коаксиальный кабель | |||
| Линия задержки | Коаксиальный кабель с подключенными клеммами | |||
| Проходные стены | Жилы в проводе эл.грамм. Три проводника | |||
| Переплетенные провода Витая пара + Инфо | Скрученные провода Скрученные провода | |||
| Телефонная линия + Инфо | Видеолиния Видеокабель | |||
| Ориентир | Исходная точка напряжения е.грамм. 5 вольт + информация | |||
| Положительный момент + Инфо | Отрицательная точка + Инфо | |||
| Терминал + Инфо | Терминал | |||
| Контрольная точка | Нейтральная точка + Инфо | |||
| Подключение переменного тока | Подключение постоянного тока | |||
| Линия приложения | Узел + информация | |||
| Конечная точка подключения Разветвление | Общая площадь | |||
| Сбалансированные клеммы | Несимметричные клеммы | |||
| Провод / гибкое соединение | Винтовой зажим Клеммные колодки + информация | |||
| Провод, гибкое соединение | Соединения клеммной колодки e.грамм. 5 подключений | |||
| Эквипотенциальность | Земля + Инфо | |||
| Эквипотенциальный + Инфо | Земля | |||
| Земля | Земля | |||
| Заземление | Паяльная перемычка + Инфо | |||
| Заземление без шума | Перемычка / перемычка + информация | |||
| Электромагнитное экранирование Закрытое экранирование + информация | Закрытая разделительная линия | |||
| Открытое экранирование | Разделительная линия | |||
| Проводящее расширение | Двойная изоляция линий | |||
| Нитиноловая проволока | Сеть низкого напряжения | |||
| Охладитель кабеля | ||||
Символы тракта передачиУсловные обозначения линий электропередачи и электрических распределительных сетейКартинная галерея линий, проводов, кабелей и электрических проводовЗагрузить символы | ||||
Структурное определение нейтрализующего эпитопа на N-концевом домене спайкового гликопротеина БВРС-КоВ
Характеристика нейтрализующего мАт 7D10, нацеленного на NTD
Для создания нейтрализующих БВРС-КоВ мАт с эпитопами вне RBD мышей иммунизировали рекомбинантный белок S БВРС-КоВ (остатки 1–1297).Затем селезенки собирали и сливали с клетками миеломы SP2 / 0, а линии гибридомных клеток проверяли на наличие положительных клонов с помощью ELISA с белком S 27 . Положительные клоны были дополнительно протестированы на их реактивность по отношению к различным фрагментам S, включая NTD субъединицы S1 (остатки 18–353), RBD (остатки 367–606) и субъединицу S2 (остатки 726–1297). Одно NTD-специфичное mAb, названное 7D10, было наконец выделено с EC 50 приблизительно 0,31 мкг / мл -1 в ELISA (рис.1а). Он не проявлял перекрестной реактивности с RBD при концентрации 4 мкг / мл -1 (рис. 1b). Мы также оценили потенциал 7D10 в форме сырых экстрактов асцита мыши для ингибирования проникновения БВРС-КоВ в чувствительные клетки Huh7 и клетки Vero E6 с псевдотипическими или инфекционными вирусами. Как и ожидалось, 7D10 был способен нейтрализовать инфекционность псевдотипированного и живого БВРС-КоВ (рис. 1c, d). Нейтрализующая активность 7D10 зависела от дозы, при этом IC 50 составлял приблизительно 0.18 мкг мл -1 против псевдотипированного вируса и практически такой же IC 50 приблизительно 0,2 мкг мл -1 против живого вируса (штамм EMC) (рис. 1c, d). Изображения, иллюстрирующие уменьшенное образование БОЕ, соответствующее скорости нейтрализации живого БВРС-КоВ, показаны на рис. 1e. Изотипирование антител показало, что 7D10 относится к подтипу IgG1. Секвенирование дополнительно определило, что сегменты V и J зародышевой линии тяжелой цепи представляют собой IGHV1-12 * 01 и IGHJ2 * 03, а сегменты легкой цепи — IGKV03-12 * 01, IGKJ1 * 01 и IGKJ1 * 02, соответственно (дополнительная таблица 1). .
Фиг. 1Специфичность связывания 7D10 и эффективность нейтрализации. Рекомбинантный NTD или RBD гликопротеина S БВРС-КоВ в концентрации 1 мкг / мл -1 использовали для покрытия планшетов в течение ночи при 4 ° C, и каждое из mAb, включая 7D10, антитело против RBD (MERS-GD27) и неродственное антитело (3C11) серийно разводили в PBS и оценивали на сродство связывания и специфичность к NTD ( a, ) и RBD ( b ). c Нейтрализация псевдотипированного БВРС-КоВ.Клетки Huh7, экспрессирующие DPP4, культивировали с 200 TCID 50 псевдотипированного MERS-CoV в присутствии серийно разведенных mAb. Процент нейтрализации рассчитывали путем измерения экспрессии люциферазы по сравнению с контрольными клетками, инфицированными псевдотипом вируса. d Нейтрализация живого БВРС-КоВ. Различные концентрации mAb предварительно культивировали с живым MERS-CoV (штамм EMC) в монослоях клеток Vero E6. Процент нейтрализации оценивали путем вычисления уменьшения количества бляшек по сравнению с инфицированным вирусом контролем.Данные представлены как среднее ± стандартное отклонение. e PFU изображения вирусной инфекции в присутствии mAb на 3 день. Изображения соответствуют процентному содержанию нейтрализации в ( d ). Примерно 30–35 PFU вирусов (штамм EMC) использовали для инфицирования клеток Vero E6 в 12-луночном планшете с mAb или без них. MERS-GD27, 3C11 mAb и PBS использовали в качестве положительного, неродственного и холостого контролей соответственно. Исходные данные представлены в виде файла исходных данных
Мы также создали химерную версию 7D10 (7D10-H), объединив V-сегменты 7D10 с основной цепью человеческого IgG1, которая была эффективно экспрессирована и очищена в клетках FreeStyle 293-F ( Дополнительный рис.1А). Эксперимент с биослойной интерферометрией (BLI) показал, что константа сродства связывания между 7D10-H и NTD составляла приблизительно 25 нМ (таблица 1 и дополнительный рисунок 1B). IC 50 очищенного 7D10-H против проникновения в клетки псевдотипированного БВРС-КоВ составлял приблизительно 0,06 мкг / мл -1 (дополнительный рисунок 1C). Мы также исследовали защитную эффективность 7D10-H против инфекции псевдотипированного MERS-CoV с использованием модели мышей R26-hDPP4 с человеческим DPP4, вставленным в локус Rosa26 с помощью CRISPR / Cas9, которые также могли быть продуктивно инфицированы MERS-CoV с высоким титром. псевдовирус, с эффектами, сопоставимыми с подлинной инфекцией 28 .Биолюминесценция репортера Fluc показала, что псевдовирусная инфекция у мышей была четко предотвращена с помощью 7D10-H и RBD-специфичных mAb MERS-4, когда оба антитела вводили внутрибрюшинной инъекцией в дозе 200 мкг на мышь (дополнительный рис. 1D). ). Рекомбинантный химерный 7D10-H, который сохранял активность мышиного 7D10 и защищал мышей R26-hDPP4 от заражения псевдотипированным MERS-CoV, использовали в последующих экспериментах по связыванию и нейтрализации.
Таблица 1 Сродство связывания 7D10-H с мутантными NTD или пезудовирусами и их нейтрализацияОбщая структура scFv 7D10, связанного с NTD
Чтобы структурно охарактеризовать 7D10 и его связывание с белком-шипом, мы определили кристаллическую структуру антитело scFv (7D10-scFv) в комплексе с NTD с разрешением 3.0 Å с конечной R рабочей 0,188 и R свободной 0,224. Статистика сбора, обработки и уточнения структуры дифракционных данных приведена в таблице 2. На каждую асимметричную единицу приходилось три комплекса 7D10-scFv, связанных с NTD. Уточненная модель содержит остатки от Tyr18 до Ser353 NTD БВРС-КоВ, от Glu1 до Ser120 из V H и от Asp26 до Lys136 из V L . N-связанные гликаны, присоединенные к Asn66, Asn104, Asn125, Asn155, Asn166, Asn222, Asn236 и Asn244 NTD, также включены в модель.Ранее было показано, что NTD MERS-CoV складывается в галектин-подобную структуру, которая может быть разделена на верхний, сердцевинный и нижний субдомены (Fig. 2a). После связывания 7D10-scFv контактирует с верхним субдоменом NTD и связанными с Asn222 гликанами со своими тяжелыми и легкими цепями (рис. 2а и дополнительный рис. 2). Все три CDR тяжелой цепи и CDR1 и CDR3 легкой цепи участвуют в связывании (фиг. 2a). Скрытая поверхность между 7D10-scFv и NTD составляет приблизительно 551 Å 2 для тяжелой цепи и 320 Å 2 для легкой цепи.
Таблица 2 Сбор данных и статистика уточнения Рис. 2Кристаллическая структура 7D10-scFv, связанная с NTD и интерфейсом связывания. a Общая структура комплекса NTD / 7D10-scFv, в котором NTD, N222-связанные гликаны на NTD, 7D10 V L и 7D10 V H окрашены в синий, серый, пурпурный и голубой цвета. , соответственно. b Эпитоп на NTD, распознаваемый 7D10. NTD представлена синей поверхностью, на которой область белка, связанная с 7D10, отображается оранжевым цветом, а гликаны, связанные с N222, отображаются в виде серых полосок. c остатка 7D10, которые участвуют в связывании. V L и V H окрашены в пурпурный и голубой цвета соответственно, а остатки, взаимодействующие с 7D10, отображаются оранжевым. d Взаимодействия между остатками 7D10 V H и соответствующими остатками NTD. e Взаимодействия между остатками 7D10 V L и соответствующими остатками NTD. f Увеличенное изображение взаимодействий между N222-связанными гликанами и 7D10
Структурные особенности интерфейса между 7D10 и NTD
Интерфейс связывания между 7D10-scFv и NTD состоит из 12 остатков и гликанов, связанных с Asn222, из NTD , а также 15 остатков из всех 6 CDR, кроме LCDR2 (рис.2б, в). Взаимодействующие остатки NTD представляют собой Tyr18, Asp20, Pro23, Asp24, Val26, Ser28, Glu188, Ser191, Asn226, Lue234, Arg235 и Asn236. Вместе с Asn22-связанными NAG508, NAG509 и MAN519 они образуют конформационный эпитоп, распознаваемый 7D10 (рис. 2b). Остатки, распознающие 7D10, представляют собой Ser31, Tyr32, Asn33 из HCDR1, Tyr52, Asn55 и Ser59 из HCDR2, Arg98, Tyr99, Asn101, Tyr102 и Tyr105 из HCDR3, Tyr59 и Tyr61 из LCDR1 и Arg2121 и Asp от LCDR3 (рис.2в). В частности, 7D10 HCDR1 остатки Ser31, Tyr32 и Asn33 взаимодействуют с Pro23 и Asp24 из NTD, и образованная водородная связь находится от 7D10 Asn33 до NTD Asp24 (рис. 2d и дополнительная таблица 2). 7D10 HCDR2 участвует в распознавании посредством Tyr52, Asn55 и Ser59 для взаимодействия Val26, Ser28 и Asn226 NTD, и две водородные связи образуются между 7D10 Tyr52 и NTD Asn226 (Fig. 2d и Supplementary Table 2). По сравнению с HCDR1 и HCDR2, HCDR3 более активно взаимодействует с NTD с его Arg98, Tyr99, Asn101, Tyr102 и Y105, взаимодействующими с Tyr18, Asp20, Pro23, Asp24 и Arg235 NTD (рис.2г). Tyr99 и Asn101 из 7D10 образуют два взаимодействия с образованием водородных связей с Asp24 NTD (дополнительная таблица 2). Для легкой цепи остатки Tyr59, Tyr61, Arg121 и Asp122 LCDR1 и LCDR3 взаимодействуют с Glu188, Ser191, Arg235 и Asn236 NTD, и между Arg121 LCDR3 и Glu188 NTD образуется солевой мостик (рис. 2e и дополнительная таблица 2). Важной особенностью интерфейса является широкое распознавание связанных с Asn222 гликанов всеми тремя CDR тяжелой цепи (фиг. 2f и дополнительная таблица 3).Специфические взаимодействия водородных связей происходят между Tyr58 и Arg98 из 7D10 и гликанов NAG508 и MAN519, соответственно (Fig. 2f и Supplementary Table 2).
Подтверждение нейтрализующего эпитопа
Чтобы подтвердить эпитоп и его критические остатки, мы провели исследование мутагенеза путем введения отдельных мутаций во все 13 распознаваемых NTD остатков, включая Trp18, Asp20, Pro23, Asp24, Val26, Ser28, Glu188, Ser191, Asn222, Asn226, Lue234, Arg235 и Asn236. Сначала мы исследовали влияние этих мутаций NTD на связывание 7D10-H.7D10-H связывал NTD дикого типа с аффинностью примерно 25 нМ (таблица 1 и дополнительный рисунок 3). Напротив, мутации D24A и R235A резко снижали связывание до уровня, который нельзя было обнаружить в эксперименте BLI (таблица 1 и дополнительный рисунок 2). Мутации E188A и N222Q снижали аффинность связывания в 148 раз до 3,7 мкМ и в 112 раз до 2,8 мкМ, соответственно (таблица 1 и дополнительный рисунок 3). Все остальные девять мутаций оказали неодинаковое влияние на связывание за счет снижения аффинности в диапазоне от 2 до 15 раз (таблица 1 и дополнительный рис.2). Эффекты этих мутаций на нейтрализующую активность 7D10-H соответствовали изменениям аффинности связывания. Псевдотипированный БВРС-КоВ, несущий мутацию D24A, E188A или R235A в спайковом гликопротеине, избежал нейтрализации 7D10-H (таблица 1 и дополнительный рисунок 4). Значения IC 50 7D10 против псевдотипированного MERS-CoV, несущего мутации D20A, V26A или N222Q, были увеличены примерно в 60, 181 и 50 раз (таблица 1 и дополнительный рисунок 4). Анализы связывания и нейтрализации в совокупности показали, что гликаны, связанные с Asp24, Val26, Glu188, Arg235 и Asn222, имеют решающее значение для распознавания и нейтрализации MERS-CoV с помощью 7D10.
7D10-H против псевдотипированного БВРС-КоВ с естественными мутациями
Секвенирование нескольких клинических изолятов показало, что гликопротеин S БВРС-КоВ развивается со средней скоростью 1,12 × 10 −3 замен на сайт в год 29 . Сопоставление депонированных последовательностей в NCBI позволило идентифицировать 22 естественных изменяющихся остатка из последовательности EMC-прототипа, включая V26F, V26I, V26A, D158Y, L411F, T424I, A482Y, L506F, D509G, V530L, V534A, E536K, D537E, V810I, Q814I, Q814 , R1020H, R1020Q, A1193S, T1202I, G1224S и V1314A, которые расположены в NTD (остатки 18–353) и RBD (остатки 367–606) субъединицы S1 и субъединице S2 (остатки 752–1297).Некоторые изменения остатков в RBD, такие как те, которые происходят в D506, D509 и E536, действительно позволили MERS-CoV избежать нейтрализации антител, нацеленных на RBD 20,21 . Учитывая, что большинство мутаций находятся за пределами NTD, мы предположили, что 7D10-H будет лучше переносить эти естественные мутации. Мы создали псевдотипированный БВРС-КоВ, несущий гликопротеины S штамма EMC, и его мутанты, несущие все 22 перечисленных изменения остатков. Анализы нейтрализации показали, что 7D10-H проявлял эффективную нейтрализующую активность против почти всех псевдотипированных вариантов БВРС-КоВ.Только две мутации V26F и V26A в NTD увеличили значение IC 50 7D10-H более чем в 150 раз и значительно снизили его нейтрализующую активность (рис. 3a, b), что подтвердило результаты структурных и биохимических исследований. исследования интерфейса привязки. Все другие встречающиеся в природе мутации, большинство из них в RBD и субъединице S2, не влияли на способность нейтрализации 7D10-H (рис. 3a, b), что указывает на то, что 7D10 будет иметь широкую широту нейтрализации против различных вариантов MERS-CoV. .
Рис. 3Ширина нейтрализации 7Д10-Н. a Нейтрализующий анализ 7D10-H против MERS-CoV дикого типа (WT) и его вариантных мутантов; сайт-направленные мутации были введены в штамм EMC для создания 22 вариантов мутантов в соответствии с естественными мутациями MERS-CoV S. b Краткое изложение опосредованного 7D10-H ингибирования инфекции всеми псевдотипными вирусами. IC 50 титры нейтрализации для мутантных вариантов EMC S представлены относительно S дикого типа (установлено на 1).Исходные данные представлены в виде файла исходных данных
Комбинация 7D10 с другими антителами, нацеленными на RBD
Все доступные в настоящее время эпитопы антител MERS-CoV с решенными структурами находятся в RBD, которые можно сгруппировать в три категории: (1) эпитоп MERS-4; (2) эпитопы MERS-27, D12, 4C2 и JC57-14; и (3) эпитопы m336, MCA1, CDC-C2 и недавно сообщенного MERS-GD27 (дополнительный рисунок 5) 25 . В нашем исследовании RBD-специфичных mAb MERS-4 мы также обнаружили синергизм с NTD-нацеливающими mAb 5F9 25 .Таким образом, выяснение эпитопа, нацеленного на 7D10, который добавил категорию вне RBD (дополнительный рисунок 5), побудило нас изучить комбинированный эффект 7D10 вместе с тремя репрезентативными антителами MERS-4, MERS-27 и MERS. -GD27 при нейтрализации псевдотипированного БВРС-КоВ путем титрования нейтрализующей активности эквимолярной смеси двух антител и сравнения реакции на дозу с реакцией, наблюдаемой в анализах нейтрализации, проводимых только с индивидуальным антителом. Как показано на рис.4, значения индекса комбинации (CI) MERS-GD27 в сочетании с 7D10 при значениях FA эффективной дозы 50%, 75%, 90% и 95% (ED 50 , ED 75 , ED 90 , и ED 95 соответственно) составляли 0,26, 0,25, 0,24 и 0,24 соответственно. Поскольку значение CI, равное 1, указывает на аддитивный эффект, <1 указывает на синергизм, а> 1 указывает на антагонизм, комбинация 7D10 и MERS-GD27 работала явно синергетическим образом. Между тем, значения индекса комбинации (CI) комбинированного MERS-4 с 7D10 при значениях FA эффективной дозы 50%, 75%, 90% и 95% (ED 50 , ED 75 , ED 90 , и ED 95 ) были 0.25, 0,27, 0,30 и 0,33 соответственно. Таким образом, комбинация MERS-4 и 7D10 также продемонстрировала синергизм, в частности, при относительно более низких концентрациях. Однако процент нейтрализации, полученный с использованием комбинированных MERS-27 и 7D10, не показал очевидной разницы в половине максимальной ингибирующей концентрации (IC 50 ) по сравнению с концентрацией только 7D10. Значения индекса комбинации (CI) комбинированных MERS-27 и 7D10 при значениях FA эффективной дозы 50%, 75%, 90% и 95% (ED 50 , ED 75 , ED 90 и ED 95 ) были 0.82, 0,87, 0,94 и 1,00 соответственно. Это указывает на то, что комбинация 7D10 с MERS-27 не проявляет ни синергизма, ни антагонизма.
Рис. 4Комбинированные эффекты для 7D10-H с RBD-специфическими mAb. a Эффекты 7D10-H в сочетании с MERS-GD27 при нейтрализации псевдотипа MERS-CoV. Процент нейтрализации рассчитывали для серийных трехкратных разведений каждого антитела отдельно и в комбинации при постоянных соотношениях в диапазоне концентраций от 27 раз до 1/81 IC 50 s.Постоянные соотношения объединенных антител составляли их IC 50 s. На оси x доза 1 была при концентрации IC 50 . Графики фракционного эффекта (FA), сгенерированные программой CompuSyn для 7D10, MERS-4 и их комбинации, демонстрируют зависимость дозировки от эффекта. График медианного эффекта рассчитанных значений CI (логарифмических) в зависимости от значений FA, на которых log CI <0 указывает на синергизм, log CI> 0 указывает на антагонизм, а log CI = 0 указывает на аддитивное действие.Процент нейтрализации, фракционный эффект и значения ДИ для 7D10 в сочетании с MERS-4 ( b ) и 7D10 в сочетании с MERS-27 ( c ) были рассчитаны и получены с использованием того же метода. Исходные данные представлены в виде файла исходных данных.
Механизм нейтрализации 7D10
Основной зарегистрированный механизм нейтрализации БВРС-КоВ основан на ингибировании связывания тримера S с клеточным рецептором DPP4. Все эпитопы этих сообщенных антител находятся в RBD, ответственной за связывание с рецептором.Тот факт, что эпитоп 7D10 находится за пределами RBD, указывает на то, что он может иметь другой механизм нейтрализации. Сначала мы проверили, способен ли 7D10 по-прежнему ингибировать связывание рецептора с помощью тримера S. FACS-анализ окрашивания клеточной поверхности показал, что scFv и Fab-фрагменты 7D10-H не ингибируют окрашивание клеток Huh7 тримером S, в то время как 7D10-H немного снижает окрашивание (рис. 5a, дополнительная таблица 4 и Дополнительный рис.6). Напротив, mAb MERS-4, нацеленное на RBD, было намного более эффективным, чем 7D10-H, в ингибировании связывания тримера S с клетками Huh7.Более того, Fab- и scFv-фрагменты MERS-4 сохраняли почти одинаковую эффективность ингибирования (фиг. 5а и дополнительная таблица 4). Анализ поверхностного плазмонного резонанса (SPR) подтвердил эти выводы, показав, что 7D10-H, а не его Fab или scFv-фрагменты, может мешать связыванию S-тримера с DPP4, связанным с чипом, дозозависимым образом (дополнительный рисунок 7). ), тогда как IgG, Fab и scFv MERS-4 все ингибировали связывание (дополнительный рис. 7).
Фиг. 57D10-H частично ингибирует связывание вируса с его рецептором DPP4. a Ингибирование связывания между рекомбинантным растворимым спайковым тримером БВРС-КоВ (S) и человеческим DPP4, экспрессируемым на поверхности клеток Huh7. Растворимый спайковый тример (S) БВРС-КоВ со стреп-меткой (1 мкг) предварительно инкубировали с моноклональными антителами (mAb) в молярном соотношении 1: 1, 1: 3, 1: 9 и 1:27 в течение 1 часа. . Клетки Huh7 инкубировали со смесями S или S и mAb в течение 1 ч. После отмывки несвязанного S клетки Huh7 окрашивали стрептавидином APC и анализировали с помощью сортировки клеток, активируемой флуоресценцией (FACS).Количество S-связанных клеток Huh-7 измеряли и характеризовали как среднюю интенсивность флуоресценции. См. Также дополнительную таблицу 4 и дополнительный рисунок 6. b — e Вид сверху и вид сбоку спайкового тримера БВРС-КоВ в активированных состояниях связывания рецептора (PDB: 5 × 5c [https://doi.org /10.2210/pdb5X5C/pdb]) с двумя RBD в верхних положениях, на которые накладываются структуры RBD / DPP4 и NTD / антитело. Тример спайка (RBD — зеленым, NTD — синим, а субъединица S2 — серым) был показан в виде рисунка.DPP4 был показан в виде полупрозрачной поверхности и окрашен в желтый цвет. Все 7D10-scFv ( b ), 7D10-Fab ( c ) и 7D10-IgG ( d , e ) были окрашены в розовый цвет. 7D10-Fab и 7D10-IgG были смоделированы с использованием гомологичного моделирования в SWISS-MODEL
. Чтобы исследовать, почему IgG, Fab и scFv из 7D10 по-разному ингибируют связывание рецептора, мы сконструировали модели их связывания с тримером S. Структуру тримера MERS-CoV S определяли с помощью крио-ЭМ с RBD в положении стоя или лежа, и только стоячий RBD мог связываться с рецептором DPP4.После наложения кристаллической структуры NTD / 7D10-scFv на тример S мы не наблюдали стерических столкновений между тремя NTD-связанными фрагментами scFv и одним или двумя RBD-связанными рецепторами DPP4 (рис. 5b и дополнительный рис. 8). Тример S с тремя рецепторами, связанными с RBD, не рассматривался, потому что крио-ЭМ исследование тримера S MERS-CoV выявило конформации только с одним или двумя стоячими RBD. Когда scFv был заменен на Fab, также не было стерических столкновений между Fab и рецептором DPP4 (рис.5в). Сложнее смоделировать связывание 7D10-H с тримером S, учитывая, что форма IgG имеет два сайта связывания и внутреннюю гибкость. Мы обнаружили, что связывание 7D10-H IgG с NTD в определенных ориентациях может ингибировать связывание DPP4 из-за стерических столкновений, в то время как стерических конфликтов с 7D10-H, связанным в некоторых других ориентациях, все еще не было (рис. 5d, e ). Эти результаты предоставили структурное объяснение неспособности 7D10-H scFv и Fab ингибировать связывание тримера S с рецептором DPP4.Они также могут объяснить, почему форма 7D10-H IgG не так эффективна, как IgG, Fab и scFv MERS-4, которые напрямую связываются с RBD.
Параллельно с биохимическими исследованиями мы также исследовали нейтрализующую активность 7D10-H IgG, Fab и scFv. Fab и scFv 7D10-H не мешали связыванию тримера S с рецептором DPP4. Однако они все еще были способны ингибировать проникновение в клетки псевдотипированного БВРС-КоВ со значением IC 50 0,26 мкг / мл -1 и 0.28 мкг мл −1 соответственно (рис. 6а). Хотя Fab и scFv 7D10-H менее активны, чем IgG в ингибировании инфекции, они все еще были сопоставимы с Fab или scFv-фрагментами нескольких известных антител, нацеленных на RBD, таких как MERS-4 Fab (IC 50 : 1,49 мкг / мл. -1 ) и MERS-4 scFv (IC 50 : 0,55 мкг мл -1 ) (дополнительный рисунок 9A). Эти результаты в совокупности указывают на то, что нейтрализация 7D10-H включает другой механизм, помимо вмешательства в начальное связывание рецептора.Мы протестировали и сравнили нейтрализующую активность 7D10 в настройках до и после прикрепления. После прикрепления клеток 7D10 все еще был способен ингибировать инфекцию псевдотипированным БВРС-КоВ с IC 50 0,55 мкг / мл -1 (фиг. 6b). Для сравнения, MERS-4, который более эффективен, чем 7D10 в ингибировании связывания рецептора, демонстрировал очень слабую нейтрализацию после связывания рецептора (дополнительный рис. 9B).
Фиг. 67D10-H, сохраняющий нейтрализующую активность после прикрепления вирусных клеток. a Кривые нейтрализации. 7D10-H IgG, 7D10-H Fab, 7D10 scFv тестировали на нейтрализующую активность против псевдотипированного БВРС-КоВ. VRC01 mAb использовали в качестве неродственного контроля. b Действия по нейтрализации привязанностей и после них. 7D10-H IgG тестировали на нейтрализующую активность против псевдотипированного БВРС-КоВ до или после связывания с рецептором. VRC01 mAb использовали в качестве неродственного контроля. c Эффект 7D10-H Fab на конформационное изменение тримерного S-тримерного БВРС-КоВ исследовали вестерн-блоттингом с использованием поликлональных антител против БВРС-КоВ S2.Рефолдинг до конформации после слияния детектировали по появлению полосы устойчивости к протеиназе-К. Трипсин использовали в количестве 5 мкг мл -1 и протеиназу К в количестве 10 мкг мл -1 . Эксперименты по перевариванию и вестерн-блоттинг проводили в трех экземплярах, и для каждого из них показан репрезентативный результат. d Разработанная нами карикатура, показывающая механизм нейтрализации, с помощью которого 7D10 блокирует проникновение БВРС-КоВ. С одной стороны, некоторые вирусные частицы не могут связываться с DPP4 из-за стерических затруднений, вызванных связыванием 7D10.С другой стороны, 7D10 все еще распознает частицы, когда up-рецептор-связывающий домен (RBD) связывается с DPP4, и может ингибировать переход субъединицы S2 от пре-слияния к пост-слиянию и инициирование слияния мембран. Исходные данные представлены в виде файла исходных данных
. Приведенные выше результаты, особенно сохранение активности 7D10 после прикрепления вируса, показали, что 7D10 также будет мешать конформационному переходу S-гликопротеина от пре-слияния к пост-слиянию, необходимого для слияния мембран.Этот переход и влияние расщепления протеазой, связывания рецептора и антител могут быть изучены биохимически путем мониторинга появления полосы, устойчивой к протеиназе-K в геле додецилсульфата натрия (SDS-PAGE), содержащем пучок шести спиралей после слияния 30, 31 . Мы показали, что гликопротеин S БВРС-КоВ в состоянии до слияния чувствителен к расщеплению протеиназы K (рис. 6c). Предыдущие исследования продемонстрировали, что расщепление по сайту S1 / S2 трипсином и связывание с клеточным рецептором значительно усиливают переход от префузии к постфузии гликопротеина шипа 31 .Соответственно, количество 50 кДа и устойчивой к протеиназе К полосы гликопротеина S, представляющей пучок из шести спиралей после слияния, было на максимальном уровне в присутствии трипсина и DPP4 (фиг. 6c). И добавление 7D10-H Fab, очевидно, уменьшило интенсивность полосы (фиг. 6c). Между тем, мы проанализировали полноразмерный тример S БВРС-КоВ, встроенный в мембрану псевдотипированного вируса, и триггером, который мы использовали для индукции конформационного перехода, была инкубация с клетками Huh 7, которые эндогенно экспрессируют рецептор DPP4.После инкубации псевдотипированного вируса с клетками Huh 7 в течение 1 часа при 37 ° C на геле SDS-PAGE появилась полоса устойчивости к протеиназе-K, и добавление 7D10-H, 7D10-H Fab или 7D10 scFv все явно уменьшало интенсивность этой полосы (дополнительный рис. 10). Таким образом, эти биохимические результаты убедительно свидетельствуют о том, что 7D10 может также проявлять свою нейтрализующую активность на стадии постприлипания после связывания с рецептором, ингибируя конформационный переход гликопротеина S, необходимый для слияния мембран (рис.6г).
Модуляция активности бактериальной сиртуиндеацилазы CobB посредством N-концевого ацетилирования
Значимость
Ацетилирование N-концевого белка у бактерий плохо изучено. Здесь мы сообщаем об идентификации Nα-ацетилтрансферазы (NAT), которая модулирует активность сиртуиндеацилазы в патогенах человека. Это важно, потому что упомянутый фермент (названный N -ацилтрансфераза A [NatA], ранее YiaC) является прокариотическим NAT не-типа Rim, а терминальное ацетилирование N — бактериального сиртуина не описано.Также важным является тот факт, что NatA влияет на метаболизм ацетата, короткоцепочечной жирной кислоты, которая, как известно, играет важную роль в патогенезе кишечника человека.
Abstract
В эукариотических клетках N-концевая аминогруппа многих белков модифицируется N-ацетилтрансферазами (NAT). Эта модификация белка может изменить укладку целевого белка; может влиять на связывающие взаимодействия целевого белка с субстратами, аллостерическими эффекторами или другими белками; или может вызвать деградацию белка.Известно, что у прокариот только рибосомные белки ацетилированы на N-конце, а ацетилтрансферазы, ответственные за эту модификацию, принадлежат к семейству белков Rim. Здесь мы сообщаем, что у Salmonella enterica длинная изоформа сиртуиндеацилазы CobB (CobB L ) ацетилируется на N-конце белком YiaC этой бактерии. Результаты анализов ацетилирования in vitro показали, что CobB L ацетилирован YiaC; Для подтверждения этих результатов использовали тандемную жидкостную хроматографию-масс-спектрометрию (LC-MS / MS).Результаты экспериментов in vitro и in vivo показали, что активность деацетилазы CobB L отрицательно влияет, когда YiaC ацетилирует свой N-конец. Мы сообщаем: 1) о модуляции активности бактериальной сиртуиндеацилазы путем ацетилирования, 2) что белок YiaC, связанный с Gcn5, является ацетилтрансферазой, которая модифицирует CobB L , и 3) что YiaC является NAT. Основываясь на наших данных, мы предлагаем название NatA ( N -ацилтрансфераза A) вместо YiaC, чтобы отразить функцию фермента.
Ацилирование белков часто встречается у прокариот и эукариот и является эффективным и быстрым средством контроля функции белка в ответ на различные стимулы (1, 2). Что выделяется в ацилировании белков, так это разнообразие органических кислот, используемых клетками для модификации белков (например, ацетат, пропионат, малонат, сукцинат и т. Д.), И большое количество ацилтрансфераз, которые катализируют модификации (1). Многие из ацилтрансфераз, которые модифицируют белки и небольшие молекулы, принадлежат к суперсемейству белков PF00583, и среди этого семейства многие белки содержат так называемый домен GNAT (связанный с GCN5 N -ацетилтрансфераза) (IPR000182).GNATs ацилируют свободные аминогруппы белков или небольших молекул (1, 2). Например, существует подмножество хорошо изученных GNAT, которые модифицируют ε-аминогруппу (Nε) в боковых цепях лизина (3, 4), в то время как другие GNAT модифицируют α-аминогруппу (Nα) исходного остатка белков (5 ⇓⇓ – 8).
В рамках работы, представленной здесь, мы отмечаем, что многие эукариотические белки ацетилированы на своих N-концах и что ацетилтрансферазы, ответственные за эти модификации, называются N-ацетилтрансферазами (NAT).Как правило, NAT катализируют перенос ацетильной группы от ацетил-кофермента A (AcCoA) к первичному амину небольшой молекулы или N-концевой аминогруппе пептида или белка. У эукариот ацетилирование Nα, как полагают, изменяет укладку белков, межбелковые взаимодействия и деградацию белков (9-12). Напротив, мало что известно о ферментах, которые катализируют ацетилирование N-концевого белка у прокариот, и о том, каковы могут быть физиологические причины такой модификации. Примерами N-концевого ацетилирования бактериальных белков, где известна ацетилтрансфераза, являются ацетилирование рибосомных белков S18, S5 и L7 / L12 ацетилтрансферазами RimI, RimJ и RimL (13–15).До недавнего времени считалось, что эти ацетилтрансферазы обладают высокой субстратной специфичностью, когда было показано, что RimI Mycobacterium tuberculosis RimI ацетилирует различные пептиды in vitro (16). Некоторые другие белки, как известно, ацетилированы Nα в бактериях, такие как шаперон секреции SecB Escherichia coli и фактор вирулентности ESAT-6 (ExsA) нескольких видов Mycobacterium . В случае белка ESAT-6 (ExsA) ацетилирование его N-конца отменяет связывающие взаимодействия с его белком-партнером CFP10 (ExsB) и снижает вирулентность Mycobacterium marinum (17⇓ – 19).Несмотря на то, что N-концевое ацетилирование у прокариот было идентифицировано, ацетилтрансферазы, ответственные за модификации, не были идентифицированы. Например, недавно опубликованные бактериальные N-концевые ацетиломы Pseudomonas aruginosa , Acinetobacter baumannii и M. tuberculosis показали, что примерно 10% белков ацетилированы на N-конце (8, 11, 20), но ферменты, ответственные за такие модификации, не идентифицированы.
Геном Salmonella enterica содержит около 26 GNAT, но функция примерно трети из них еще не определена.Кроме того, геном S. enterica содержит одну никотинамидадениндинуклеотид (NAD + ) -зависимую сиртуиндеацилазу CobB, функция которой работает совместно с ферментом протеинацетилтрансфераза (Pat), обратимо модулируя активность ацетил-CoA (Acs) (21, 22). Как показано на схеме 1, реакция CobB-зависимого деацетилирования потребляет NAD + и дает O — ацетил-ADP (аденозиндифосфат рибоза) и никотинамид.
Интересно, что эта бактерия синтезирует две биологически активные изоформы CobB, называемые как CobB S (короткая изоформа CobB) и CobB L (длинная изоформа CobB), которые отличаются по размеру N-концевым удлинением каталитического ядра из 37 остатков (рис.1).
Рис. 1.Биологически активные изоформы сиртуиндеацилазы CobB S. enterica . N-концевая аминокислотная последовательность CobBL. Остатки, выделенные желтым цветом, представляют собой гидрофобные аминокислоты; красные остатки — аргинины, синие — лизины.
Наша группа показала, что обе изоформы CobB деацетилировали свой истинный белковый субстрат (т.е. ацетилированную ацетил-CoA синтетазу [Acs Ac ]) in vivo и in vitro (23). Однако физиологическая значимость двух изоформ CobB в S.enterica неизвестно. Здесь мы сообщаем, что изоформа сиртуиндеацилазы CobB L этой бактерии ацетилирована на N-конце и что предполагаемая ацетилтрансфераза YiaC ацетилирует N-конец CobB L . Мы также сообщаем о доказательствах in vivo и in vitro, что YiaC-зависимое N-концевое ацетилирование CobB L отрицательно влияет на его деацетилазную активность.
Результаты
YiaC Ацетилаты CobB
L , но не CobB S .Поиск белковых субстратов для S.enterica предполагаемые GNATs привели нас к открытию, что белок S. enterica YiaC ацетилировал S. enterica CobB L , но не CobB S . Как показано на фиг. 2, когда обе изоформы S. enterica CobB инкубировали с [1- 14 C] -AcCoA в зависимости от YiaC, радиоактивная метка переносилась на CobB L , но не на CobB S. (рис. 2, дорожки 4 и 5 соответственно). Поскольку CobB S не был ацетилирован, мы предположили, что сайты ацетилирования расположены в пределах 37-аминокислотного N-концевого, богатого аргинином мотива CobB L (рис.1).
Рис. 2.YiaC ацетилирует CobB L , но не CobB S . YiaC-зависимое ацетилирование изоформ CobB оценивали после инкубации белков с [1- 14 C] -AcCoA (20 мкМ) в течение 1 ч при 37 ° C. Подробные условия анализа описаны в Materials and Methods . Контроли включали инкубацию изоформ CobB с [1- 14 C] -AcCoA в отсутствие YiaC. Белки разделяли с помощью SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) и визуализировали окрашиванием Coomassie Brilliant Blue R ( Upper ) с использованием стандартов белка Precision Plus (Bio-Rad) в качестве маркеров молекулярной массы (дорожка 1, MM, килодальтон) .Сигнал радиоактивной метки визуализировали с помощью фосфорного изображения ( Нижний ). * Радиоактивно меченые ацетильные группы.
YiaC не ацетилирует Nε-аминогруппы остатков лизина.
N-концевое удлинение CobB L содержит два лизина (K14, K16), которые мы исследовали как возможные сайты ацетилирования. Чтобы проверить эту возможность, мы заменили K14 и K16 на аланин независимо и в комбинации. Три варианта (CobB L K14A , CobB L K16A и CobB L K14A, K16A ) были произведены в избыточном количестве, изолированы и инкубированы с YiaC в присутствии [1707–]. C] -AcCoA.Неожиданно YiaC ацетилировал все три варианта CobB L (фиг. 3, дорожки 5, 7 и 9), что позволяет предположить, что в используемых условиях анализа YiaC не изменяет положение Nε ни K14, ни K16. Отметим, что интенсивность сигнала для ацетилированного варианта CobB L K14A, K16A была меньше, чем для вариантов с одной аминокислотой, и что интенсивность сигнала была соизмерима с количеством белка, загруженного на гель. Выход варианта CobB L K14A, K16A был ниже, чем у вариантов с одной аминокислотой.
Рис. 3.YiaC не ацетилирует лизильные остатки CobB L . Для запроса сайта модификации лизиновые варианты CobB L очищали и инкубировали с YiaC и [1- 14 C] -AcCoA. Цель состояла в том, чтобы определить, является ли YiaC Nα-ацетилтрансферазой. Этот эксперимент был проведен, как описано для рис. 2, где варианты CobB L (3 мкМ, за исключением CobB L K14A, K16A был при 2 мкМ) инкубировали с [1- 14 C] -AcCoA и либо с или без YiaC (1 мкМ).Дорожки 4 и 5 содержат CobB L K14A , дорожки 6 и 7 содержат CobB L K16A , а дорожки 8 и 9 содержат CobB L K14A, K16A . Результаты контрольных экспериментов с использованием CobB L плюс AcCoA с YiaC или без него показаны на дорожках 2 и 3 соответственно. Дорожка 1 представляет маркер молекулярной массы (ММ) в килодальтонах. SDS-PAGE, электрофорез в полиакриламидном геле с додецилсульфатом натрия. * Радиоактивно меченые ацетильные группы.
YiaC изменяет N-конец CobB
L In vitro.Для подтверждения нашей гипотезы о том, что CobB L является ацетилированным Nα, была проведена жидкостная хроматография-тандемная масс-спектрометрия (ЖХ-МС / МС). Результаты анализа пептидного фингерпринта ацетилированной длинной изоформы CobB ( Ac CobB L ) однозначно показали, что N-конец CobB L ацетилирован YiaC (фиг. 4).
Рис. 4.Масс-спектрометрический анализ Ac CobB L . CobB L (5 мкМ) инкубировали с AcCoA (1 мМ) с YiaC (3 мкМ) и без него при 37 ° C в течение 1 ч.Компоненты реакционной смеси разделяли с помощью SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия). CobB L вырезали из геля и отправляли в центр масс-спектрометрии Биотехнологического центра Университета Висконсина в Мэдисоне для анализа ЖХ-МС / МС (жидкостная хроматография-тандемная масс-спектрометрия). ( A ) Хроматографическая кривая для выбранного интересующего иона ( Ac GTMQSR). EIC, хроматограмма экстрагированных ионов. ( B ) Масс-спектр Ac CobB L . b Ионы представляют собой серию фрагментов, которые отходят от N-конца; Ионы и представляют собой серию фрагментов, которые отходят от С-конца. Ось x для m / z обозначает массу (m) над числом заряженных ионов (z). Программа MASCOT (http://www.matrixscience.com) была поисковой машиной в Интернете, используемой для идентификации пептидов на основе их массы.
Для подтверждения данных ЖХ-МС / МС были синтезированы два пептида из первых 50 аминокислот CobB L (Пептид 2.0, Вирджиния): один начинался с немодифицированного l-Met, а второй — с l-Met Ac . Анализы ацетилирования in vitro проводили с указанными выше пептидами в качестве субстратов для YiaC. Когда к реакционной смеси добавляли [1- 14 C] -AcCoA, YiaC ацетилировал пептиды CobB L , N-концевая аминогруппа которых не была модифицирована (фиг. 5, дорожки 2 и 8), но не ацетилировала пептиды. Пептид CobB L , первым остатком которого был l-Met Ac (фиг. 5, дорожка 5).В совокупности эти данные показали, что YiaC является протеинацетилтрансферазой Nα (NAT).
Рис. 5.YiaC ацетилирует N-концевой метионин CobB L . SDS-PAGE и анализ фосфорной визуализации неацетилированных или ацетилированных на N-конце синтетических пептидов аминокислот, охватывающих первые 50 остатков CobB L , которые инкубировали с YiaC без [1- 14 C] -AcCoA (дорожки 3 и 6 ), с [1- 14 C] -AcCoA, но без YiaC (дорожки 4 и 7), или с YiaC и [1- 14 C] -AcCoA (дорожки 2, 5 и 8).Дорожки 1 и 9 представляют собой стандартные маркеры молекулярной массы (ММ) белка Precision Plus (килодальтон). SDS-PAGE, электрофорез в полиакриламидном геле с додецилсульфатом натрия. * Радиоактивно меченые ацетильные группы.
N-конец
Ac CobB L Не может быть деацетилирован ни одной изоформ CobB.Чтобы проверить, может ли Ac CobB деацетилироваться с N-конца любой изоформой CobB, были проведены анализы ацетилирования с помощью YiaC, CobB L и [1- 14 C] -AcCoA.Меченные радиоактивным изотопом ацетилированные длинные изоформы CobB (* Ac CobB L ) освобождали от избытка AcCoA и инкубировали либо с CobB L , либо с CobB S , с или без NAD + . Положительные контроли также были добавлены, чтобы показать, что CobB L и CobB S деацетилировали свой истинный субстрат, Acs Ac . Как видно на фиг.6, CobB L и CobB S деацетилированная радиоактивно меченная ацетил-CoA синтетаза (Acs * Ac ; дорожки 3 и 5), но не деацетилировала * Ac CobB L дорожки 7 и 9).Это показало, что N-концевое ацетилирование CobB L не было обращено ни одной изоформ CobB.
Рис. 6.Nα ацетилированные белки не являются субстратами для CobB. [1- 14 C] — Ac CobB L инкубировали либо с CobB L , либо с CobB S с (дорожки 7 и 9) и без NAD + (дорожки 6 и 8). Положительные контроли немеченых CobB L и CobB S также тестировали на их способность деацетилировать [1- 14 C] -Acs Ac (дорожки 2-5).Дорожка 1 представляет стандарты молекулярной массы (ММ), выраженные в килодальтонах. Образцы разделяли с помощью SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), и перенос метки [1- 14 C] выявляли с помощью анализа изображения люминофора. * Радиоактивно меченые ацетильные группы.
Доказательства in vitro и in vivo, что Nα-ацетилирование CobB
L изменяет его деацетилазную активность.Доказательства in vitro.
N-концевое ацетилирование CobB L с помощью YiaC (рис.2) вызвали вопросы относительно влияния модификации на ферментативную активность CobB L . Чтобы ответить на этот вопрос, мы провели анализы деацетилирования белка in vitro с Acs * Ac и Ac CobB L , чтобы улучшить нашу способность обнаруживать события деацетилирования. Протоколы для создания Acs * Ac и * Ac CobB L описаны в «Материалы и методы» . После того, как у нас были Acs * Ac и * Ac CobB L , был проведен следующий эксперимент.CobB L или * Ac CobB L добавляли к реакционной смеси, содержащей Acs * Ac и NAD + , и образцы (по 25 мкл каждый) инкубировали в течение 1 часа. Как видно на фиг.7 A , неацетилированный CobB L деацетилированный Acs * Ac , на что указывает исчезновение сигнала на изображении люминофора, соответствующего Acs * Ac (молекулярная масса ∼72 кДа) (рис. 7 А , переулок 5). Напротив, * Ac CobB L не деацетилировал Acs * Ac так же эффективно, как CobB L (рис.7 A , дорожка 3 по сравнению с полосой 5). Количественная денситометрия сигналов исчезновения радиоактивной метки Acs Ac показала в среднем ~ 50% снижение активности деацетилазы CobB L при ацетилировании. Результаты, показанные на рис. 7, представляют собой серию из шести отдельных экспериментов. Эти данные показали, что модификация отрицательно повлияла на ферментативную активность * Ac CobB L .
Рис. 7. Ацетилирование N-конца CobB L отрицательно влияет на его деацетилазную активность in vitro и in vivo.( A ) Для оценки ферментативной активности Ac CobB L in vitro, белок Acs, радиоактивно меченный [1- 14 C] -AcCoA (Acs Ac *), инкубировали либо с CobB L . (дорожка 5) или * Ac CobB L (дорожка 3) и NAD + для визуализации удаления радиоактивно меченной ацетильной группы из Acs. CobB L ацетилировали YiaC, используя [1- 14 C] -AcCoA в качестве субстрата. * Ac CobB L помог визуализировать подвижность Ac CobB L на изображении люминофора.Отрицательные контроли включали реакции, перечисленные выше, за исключением добавления NAD + (дорожки 2 и 4). Дорожка 1 показывает маркер молекулярной массы (ММ) в килодальтонах. SDS-PAGE, электрофорез в полиакриламидном геле с додецилсульфатом натрия. ( B ) Все штаммы выращивали на минимальной среде NCE (незаменимая без углерода) с добавлением ацетата (10 мМ) в качестве единственного источника углерода и энергии. Используемые штаммы: Δ cobB / pCV1 / pCV3 (черные треугольники; отрицательный контроль), Δ cobB / p cobB + (CobB S ) / pCV1 (пустые квадраты; комплементация), Δ cob / p cobB + (CobB L ) / pCV1 (черные квадраты; дополнение), cobB + / pCV1 / pCV3 (серые квадраты; контроль дикого типа), cobB / p yiaC + / pCV3 (светлые кружки), Δ cobB / p cobB + (CobB S ) / p yiaC + (открытые треугольники ) и Δb / p cobB + (CobB L ) / p yiaC + (серые круги).Клонирующие векторы pCV1 и pCV3 содержат один и тот же индуцируемый арабинозой промотор (24, 25). Карты клонирующих векторов pCV1 и pCV3 можно найти в ссылке. 25. Используемая концентрация арабинозы составляла 100 мкМ. Каждый штамм выращивали в трех биологических и технических повторностях, и анализы проводили три независимых раза. Планки погрешностей представляют стандартное отклонение технических трех повторностей. * Радиоактивно меченые ацетильные группы. Чтобы убедиться, что ацетилирование N-конца CobB L влияет на его ферментативную активность, мы провели эксперименты in vivo для оценки активности CobB L как функции YiaC во время роста на минимальной среде, содержащей низкую концентрацию ацетата. (10 мМ).Известно, что в таких условиях функция CobB необходима для поддержания деацетилированных Acs, следовательно, активных (3, 22, 26). Для этого мы переместили две плазмиды в штамм S. enterica Δ cobB . Одна плазмида кодировала либо CobB L (M37A, M38A) , либо CobB S (M1A) ; вторая плазмида кодировалась YiaC или векторным контролем. Гены cobB и yiaC находились под контролем l — (+) — арабинозы-индуцибельного промотора P araBAD (24).Чтобы получить только CobB L и не белок CobB S , плазмида, кодирующая CobB L , содержала естественный исходный метионин (M1) для CobB L , но имела две мутации, которые изменяют исходный метионин и соседний метионин на CobB . S (т.е. M37 и M38) на аланины. Аллель cobB , кодирующий CobB M37A, M38A , эффективно блокирует синтез CobB S (23). И наоборот, чтобы клетка могла синтезировать только CobB S , плазмида, кодирующая CobB S , имеет аллель cobB с первым метионином, измененным для кодирования аланина, так что белок CobB L не может быть получен из M1, что приводит к эксклюзивный синтез CobB S , начиная с позиции M38 (23).Поведение роста штаммов Δ cobB и cobB + , несущих пустые клонирующие векторы, использовали в качестве контроля. В качестве дополнительного контроля использовали штамм cobB + , несущий плазмиду, несущую yiaC + (светлые кружки на фиг. 7 B ). Все штаммы выращивали на минимальной среде с ацетатом (10 мМ) в качестве единственного источника углерода и энергии. Данные, представленные на фиг. 7 B , показывают поведение роста представляющих интерес штаммов в присутствии индуктора [1 — (+) — арабинозы, 100 мкМ].Были сделаны следующие наблюдения. 1) Как и ожидалось, штамм Δ cobB не смог вырасти на 10 мМ ацетате (черные треугольники на фиг. 7 B ), потому что Acs оставались ацетилированными, следовательно, неактивными. 2) Фенотип штамма Δ cobB был скорректирован эктопической экспрессией аллелей cobB , которые управляли синтезом функционального CobB S (белые квадраты на фиг.7 B ) или CobB L (черные квадраты) на рис.7 B ). 3) Синтез YiaC не повлиял на рост штамма cobB + (белые кружки vs.серые квадраты на рис.7 ( B ). 4) Синтез CobB S и YiaC штаммом с геномной делецией cobB привел к поведению роста, которое было очень похоже на поведение штамма cobB + , который синтезировал YiaC (открытые треугольники против серого квадраты и белые кружки на рис.7 ( B ). 5) Синтез CobB L и YiaC штаммом Δ cobB преждевременно остановил рост (серые кружки на фиг. 7 B ). Эти результаты согласуются с нашими данными in vitro, которые показали, что N-концевое ацетилирование CobB L с помощью YiaC отрицательно влияет на деацетилазную активность CobB L (рис.7 А ). Опять же, в используемых условиях роста снижение активности деацетилазы может предотвратить деацетилирование (следовательно, реактивацию) Acs, блокируя превращение ацетата в AcCoA с сопутствующим отрицательным эффектом на рост клеток. Как предсказано вышеупомянутыми результатами, удельная активность Acs различалась между штаммом Δ cobB , который эктопически синтезировал CobB L и YiaC, и штаммом Δ cobB , который синтезировал CobB L , но не синтезировал эктопически. YiaC (рис.8 А ). В вышеупомянутых штаммах мы измерили статистически значимое снижение активности Acs при избыточном продуцировании YiaC. Эти результаты согласуются с данными, показывающими, что YiaC-зависимое ацетилирование N-конца CobB L снижает его активность, поддерживая ацетилирование Acs (следовательно, неактивное) и, таким образом, останавливая рост из-за пониженных уровней AcCoA. Acs снижается, когда YiaC чрезмерно продуцируется в S. enterica . ( A ) Штаммы S.enterica с удаленным cobB и только с pCobB L M37A, M38A в транс с или без pYiaC выращивали до мидлогарифмической фазы с 10 мМ ацетатом в качестве единственного источника углерода и энергии, лизировали и тестировали на Acs-специфические активность от 4 мкг лизата. Удельную активность (микромоль AMP [аденозинмонофосфат] минута -1 миллиграмм -1 ) рассчитывали с использованием непрерывного спектрофотометрического анализа, описанного в Материалы и методы .Активность Acs снижалась в лизатах штамма с гиперэкспрессией pYiaC. Эксперимент проводили в трех биологических повторах по девять технических повторов в каждом. * P = 0,03. ( B ) Концентрация белка Acs одинакова в обоих штаммах, испытанных на активность Acs, на основании количественного анализа вестерн-блоттинга лизатов, использованных в эксперименте в A , на анти-Acs. Планки погрешностей представляют непарный тест t с одинаковым стандартным отклонением; не значимо (нс) P значение = 0.5118. Для проверки того, что концентрация Acs была одинаковой в обоих штаммах, проводили количественный вестерн-блоттинг с использованием кроличьих поликлональных анти-Acs-антител для определения количества белка Acs, присутствующего в лизатах, используемых для анализа активности Acs. Действительно, лизаты штаммов, которые либо сверхэкспрессировали yiaC + , либо несли пустой вектор, содержали такое же количество Acs (фиг. 8 B и SI, приложение , фиг. S2). У эукариот некоторые ацетилированные на N-конце белки подвергаются деградации (27). Другие примеры показывают, что N-концевое ацетилирование стабилизирует ацетилированные белки и предотвращает деградацию (28, 29). Чтобы оценить, изменило ли ацетилирование CobB L уровни белка CobB L in vivo, мы использовали кроличьи поликлональные антитела против CobB для количественного определения уровней CobB в бесклеточных экстрактах трех штаммов: yiaC + / вектор, yiaC :: kan + / vector и yiaC :: kan + / pYiaC.Штаммы выращивали в минимальной среде с добавлением ацетата (10 мМ) и 1 — (+) — арабинозы (500 мкМ) или в богатой среде (Lysogeny Broth, LB), содержащей 1 — (+) — арабинозу (1 мМ). В обоих случаях рост отслеживали при 600 нм. Данные, представленные на фиг. 9, показывают, что во время роста на богатой среде или на минимальной среде с низкой концентрацией ацетата концентрация CobB L не изменялась ни у одного из штаммов. Чистые белковые контроли CobB L и CobB S были добавлены в качестве положительных контролей, и был включен блот анти-DnaK, чтобы гарантировать, что все образцы были загружены одинаково.Статистический анализ плотности пикселей вестерн-блотов анти-CobB показывает отсутствие разницы в концентрации CobB L ни в одном из протестированных штаммов, а также отсутствие разницы в CobB S или белке внутреннего контроля DnaK (фиг. 9 C и SI Приложение , рис. S5). YiaC не изменяет уровни белка CobB in vivo. Концентрацию белка изоформы CobB в штаммах S. enterica , в которых yiaC был удален или дополнен pYiaC, сравнивали с исходным штаммом ( yiaC + / вектор) с помощью вестерн-блоттинга.( A ) Вестерн-блоттинг клеток, выращенных на среде LB с 1 мМ l — (+) — арабинозы с использованием антител против CobB или против DnaK. Клетки собирали при трех различных оптических плотностях (OD 600 нанометров, нм) 0,2, 0,5 и 0,7. ( B ) Вестерн-блоттинг уровней белка CobB или уровней белка DnaK из клеток, выращенных с 10 мМ ацетатной минимальной средой с добавлением 500 мкМ l — (+) — арабинозы. Вестерн-блоттинг против DnaK использовали в качестве положительного контроля, чтобы показать, что все дорожки были загружены одинаково, и использовать в качестве стандарта для расчетов денситометрии.( C ) Плотность пикселей для образца только CobB L вестерн-блоттинга против CobB из B , показывающая, что концентрация CobB L во всех штаммах одинакова. Расчеты проводились с использованием однофакторного дисперсионного анализа, и различия концентраций белка CobB L не являются значимыми при значении P , равном 0,9747. В S. enterica белок YiaC представляет собой ацетилтрансферазу Nα, которая контролирует активность длинной изоформы сиртуиндеацилазы CobB.Представленные здесь данные подтверждают несколько выводов относительно функции белка YiaC S. enterica . Во-первых, YiaC обладает активностью Nα-ацетилтрансферазы. Наши данные также показывают, что YiaC модифицирует длинную изоформу сиртуиндеацилазы CobB (CobB L ) этой бактерии, но не короткую изоформу CobB (рис. 2). Эти результаты поместили сайты ацетилирования внутри 37-аминокислотного N-концевого удлинения CobB L (фиг. 1). Во-вторых, YiaC не модифицирует Nε-аминогруппы лизиновых боковых цепей белка CobB L (рис.3 и 4). Может ли YiaC модифицировать лизильные остатки других белков, еще предстоит определить. В-третьих, YiaC, по-видимому, обладает довольно широкой специфичностью в отношении своей мишени, поскольку он ацетилирует аминогруппу Nα глицилового остатка, который остается слитым с белком после обработки протеазой для удаления метки MBP (мальтозосвязывающий белок) -H 6 (рис. 2 и 4). YiaC также ацетилирует Nα-аминогруппу N-концевого метионина CobB L с С-концом H 6 , и N-концевой метионин синтетического пептида содержит первые 50 аминокислот CobB L . (Рисунок.5 и SI Приложение , рис. S4). Мы отмечаем, что неоднократные попытки выделить Ac CobB L на N-конце из клеток были безуспешными, несмотря на то, что несколько различных протеаз использовали в комбинации до масс-спектрометрического анализа. Мы полагаем, что аминокислотный состав N-конца CobB L затрудняет этот анализ, поскольку он очень богат аргининами. Однако, когда мы удалили N-концевую метку из MBP-H 6 -CobB L , полученный белок имел два дополнительных остатка на своем N-конце, а именно Gly-Thr (GT-CobB L ).YiaC ацетилировал GT-CobB L in vitro (рис. 4 и SI, приложение , рис. S4), но не ацетилировал белок, который имел N-концевую гексагистидиновую метку (белок H 6 -CobB L ) ( SI Приложение , рис. S4). В совокупности наши результаты показывают, что YiaC является Nα-ацетилтрансферазой (NAT) и что изоформа CobB L является ее субстратом. Насколько нам известно, YiaC — это бактериальный NAT, который не принадлежит к семейству белков Rim.Основываясь на наших данных, мы предлагаем изменить название YiaC на NatA, чтобы отразить тот факт, что это Nα-ацетилтрансфераза. Недавно Christensen et al. (30) показали, что сверхэкспрессия yiaC в штамме с делецией E. coli pta patZ acs cobB демонстрирует повышенное ацетилирование белка, измеренное с помощью вестерн-блоттинга с использованием антител α-AcK. Эти результаты интересны, потому что эти исследователи провели обогащение AcK и масс-спектрометрию, чтобы идентифицировать предполагаемые мишени лизина белка YiaC.CobB не был идентифицирован из вышеупомянутого протеома, потому что ген cobB был удален в используемых штаммах, а N-концевое ацетилирование белков не сообщалось. Как упоминалось выше, в настоящее время мы не можем исключить возможность того, что YiaC ацетилирует лизильные остатки, как было предложено Christensen et al. (30). Необходима дополнительная работа, чтобы определить, может ли YiaC выполнять ацетилирование белков Nα и Nε. Выявленная активность YiaC как N-концевой ацетилтрансферазы поднимает вопросы о роли N-концевого ацетилирования в физиологии прокариотических клеток. Мы показали, что N-концевое ацетилирование CobB L происходит как in vitro, так и in vivo, а также что YiaC-зависимое ацетилирование CobB L отрицательно влияет на его деацетилазную активность, которая затем отрицательно влияет на рост ацетата. Каким образом добавление ацетильной группы к N-концу CobB L влияет на его активность, представляет интерес, и текущие исследования в нашей лаборатории сосредоточены на том, чтобы ответить на этот вопрос.YiaC — это NAT, который не принадлежит к семейству белков Rim ацетилтрансфераз Nα с аминогруппой. Также мы сообщаем об ацилировании прокариотической сиртуиндеацилазы. Как указывалось выше, на основании представленных здесь данных мы предлагаем изменить название YiaC на NatA (для Nα-ацетилтрансферазы), чтобы отразить биохимическую активность фермента. Подробные протоколы, использованные в этом исследовании, представлены в приложении SI . SI Приложение содержит протоколы очистки белков CobB S , CobB L и YiaC; эксклюзионная хроматография; анализы ацетилирования и деацетилирования in vitro; протокол определения активности Acs in vitro; количественный вестерн-блот-анализ; списки штаммов, плазмид и праймеров; методы конструирования штаммов и плазмид; питательные среды, условия роста и анализ поведения роста; и масс-спектрометрический анализ состояния ацетилирования представляющих интерес белков.Приложение SI также содержит таблицы, которые содержат информацию о штаммах, плазмидах и праймерах, а также гомологах YiaC и CobB в других кишечных бактериях, соответственно ( SI Приложение , таблицы S1 – S4). Кроме того, приложение SI содержит рисунки (приложение SI , рисунки S1 – S5), которые представляют результаты контрольных экспериментов, данные относительно олигомерного состояния YiaC в растворе и количественную оценку вестерн-блотов изоформ CobB или DnaK (контроль ), использованный в этом исследовании. Все данные этой работы приведены в статье. Мы благодарим Гжегожа Сабата из Биотехнологического центра Университета Висконсин-Мэдисон за выполнение анализа ЖХ-МС / МС. Рэйчел Буркхардт первой наблюдала ацетилирование CobB. Эта работа была поддержана грантом NIH R35 GM130399 (J.C.E.-S.). Автор: A.R.P. и J.C.E.-S. спланированное исследование; А.Р.П. провела исследование; A.R.P. и J.C.E.-S. проанализированные данные; J.C.E.-S. концептуализировал проект; и A.R.P. и J.C.E.-S. написал газету. Авторы заявляют об отсутствии конкурирующей заинтересованности. Эта статья представляет собой прямую публикацию PNAS. Эта статья содержит вспомогательную информацию в Интернете по адресу https://www.pnas.org/lookup/suppl/doi:10.1073/pnas.2005296117/-/DCSupplemental. Специфический RIA был разработан для измерения N-концевой иммунореактивности предсердного натрийуретического фактора (ANF) в плазме крови человека.Антитела кроликов против N-концевого фрагмента ANF крысы [ANF- (11–37)] имели 100% перекрестную реактивность с ANF- (1–30) человека и очищенной N-концевой иммунореактивностью ANF в плазме. ED 80 и ED 50 стандартных кривых, полученных с использованием [ 125 I] человеческого ANF- (1–30) и человеческого ANF- (1–30), составили 31,5 ± 5,4 (± стандартное отклонение) и 132,5 ± 20,4 фмоль / туба соответственно. Концентрации N-концевого пептида ANF в плазме определяли напрямую, без экстракции, поскольку разбавление плазмы и добавление стандарта к плазме приводили к параллельным дозозависимым ответам в RIA и практически 100% извлечению ANF- (1–30), добавленного в плазму.Очистка N-концевой иммунореактивности ANF из 1,5 л человеческой плазмы с помощью аффинной хроматографии и аминокислотного секвенирования позволила предположить, что она была тесно связана с ANF- (1–98), хотя также были обнаружены некоторые деградированные пептиды. Средний базальный уровень N-концевого пептида ANF в плазме, измеренный у 34 здоровых субъектов, составил 420 ± 157 (± стандартное отклонение) пмоль / л. Значения были выше в плазме пациентов с застойной сердечной недостаточностью (степени III и IV; 7 041 ± 6 136 пмоль / л; n = 13) или хронической почечной недостаточностью (10 079 ± 4942 пмоль / л; n = 20).У 9 пациентов с хронической почечной недостаточностью гемодиализ привел к снижению уровня ANF- (99–126) в плазме на 30% ( P <0,05) с 34,7 ± 12,3 (± стандартное отклонение) до 23,2 ± 6,1 пмоль / л, но отсутствие изменений концентрации N-концевого пептида ANF в плазме. Эти данные указывают на то, что N-концевая часть про-ANF секретируется с ANF- (99–126). Его более высокие уровни в плазме в базальном состоянии и при хронической почечной недостаточности предполагают другой процесс выведения, чем у ANF- (99–126), который может частично опосредоваться почками. Этот контент доступен только в формате PDF. Цель . Сравнить концентрации N-концевого про-C-натрийуретического пептида в плазме у беременных с нормальным давлением, пациентов с легкой преэклампсией и пациентов с тяжелой преэклампсией. Методы . Мы собрали образцы венозной крови у 25 беременных с нормальным АД, у 15 пациентов с преэклампсией легкой степени и у 15 пациентов с преэклампсией тяжелой степени.Женщины находились на сроке от 30 до 40 недель беременности и в возрастном диапазоне от 20 до 35 лет. Уровни N-концевого натрийуретического пептида про-C-типа измеряли с помощью ELISA. Статистические сравнения были выполнены с помощью одностороннего дисперсионного анализа, тестов Краскела – Уоллиса и Манна – Уитни U . Результатов . Средние значения (межквартильный размах — IQR) N-концевого натрийуретического пептида про-C-типа составляли 6,48 (3,33) пмоль / л в группе женщин с нормальным АД, 7,37 (3,43) пмоль / л у пациентов с легкой преэклампсией и 11 пмоль / л. .52 (6,13) пмоль / л у пациентов с тяжелой преэклампсией. Уровень N-концевого натрийуретического пептида про-C-типа был значительно повышен в группе исследования тяжелой преэклампсии (), тогда как не было существенной разницы между группами с легкой преэклампсией и группами с нормальным давлением (). Заключение . Наши данные показывают, что концентрация в плазме N-концевого натрийуретического пептида про-C-типа значительно повышена у пациентов с тяжелой преэклампсией, но не у пациентов с легкой преэклампсией.Тяжесть преэклампсии может быть связана с циркулирующими уровнями концентраций N-концевого натрийуретического пептида про-С-типа. Семейство натрийуретических пептидов состоит из трех основных пептидов, имеющих одинаковую кольцевую структуру из 17 аминокислот: натрийуретический пептид A-типа (предсердный), натрийуретический пептид B-типа (мозг) и натрийуретический пептид C-типа. [1]. Эти натрийуретические пептиды формируют свои биологические эффекты посредством активации трех различных рецепторов натрийуретических пептидов, которые ранее были описаны как рецепторы натрийуретических пептидов A, B и C [2].Натрийуретический пептид A-типа и натрийуретический пептид B-типа вызывают свои биологические эффекты через рецептор натрийуретического пептида A, тогда как натрийуретический пептид C-типа действует более избирательно с рецептором натрийуретического пептида B. Оба этих рецептора натрийуретического пептида увеличивают внутриклеточный уровни циклического гуанозинмонофосфата, что приводит к нескольким различным физиологическим изменениям, влияющим на различные гомеостатические процессы [3]. В отличие от рецепторов натрийуретического пептида А и В, рецептор натрийуретического пептида С действует как рецептор клиренса.Помимо очищающей функции, он также оказывает важное влияние на тонус кровеносных сосудов [4]. Натрийуретический пептид С-типа был третьим идентифицированным членом этого семейства. Натрийуретический пептид С-типа представляет собой пептид местного действия аутокринным / паракринным образом. Он присутствует в плазме в очень низких концентрациях вследствие его быстрого выведения из кровотока [5]. Он обладает широким спектром биологических действий, таких как расширение сосудов, ингибирование роста гладкомышечных клеток сосудов [6], модуляция активности иммунных клеток и тромбоцитов [7] и ремоделирование сосудов.Кроме того, повышенные уровни натрийуретического пептида С-типа в плазме обнаруживаются у пациентов с септическим шоком, сердечной недостаточностью и почечной дисфункцией, что связывает этот пептид, полученный из эндотелия, с сердечно-сосудистыми и эндотелиальными патологиями [2]. Преэклампсия и эклампсия — два гипертонических расстройства во время беременности. Патофизиологические механизмы в развитии этих нарушений еще не ясны, хотя хорошо известно, что эндотелиальная дисфункция играет фундаментальную роль [8].Насколько нам известно, в литературе есть две публикации, в которых сравниваются уровни натрийуретического пептида С-типа у пациентов с преэклампсией и здоровых беременных женщин с нормальным давлением. В одном из этих исследований уровни натрийуретического пептида С-типа в плазме определялись с помощью иммуноферментного анализа (ELISA), и не было обнаружено никаких существенных различий [9]. В другом исследовании оценивали уровни в плазме N-концевого натрийуретического пептида про-С-типа (пептид-предшественник натрийуретического пептида С-типа с более длительным периодом полужизни), и Prickett et al.обнаружили, что уровни в плазме N-концевого натрийуретического пептида про-C-типа были выше у пациентов с преэклампсией [10]. В данном исследовании мы стремились измерить уровни в плазме N-концевого натрийуретического пептида про-C-типа у пациентов с преэклампсией и проанализировать его связь с тяжестью преэклампсии. Всего в этом исследовании приняли участие 55 беременных женщин, наблюдаемых в отделении акушерства и гинекологии Образовательно-исследовательской больницы по женским и детским болезням Зейнеп Камил, Турция.В группу преэклампсии были отобраны 15 пациентов с тяжелой преэклампсией и 15 пациентов с легкой преэклампсией. Письменное информированное согласие было получено от всех женщин. Все отобранные женщины были в возрасте от 20 до 35 лет и имели одноплодную беременность. Пациенты с диабетом, сердечными или почечными заболеваниями и хронической гипертонией были исключены из исследования. Также из исследования были исключены женщины с диагнозом HELLP-синдром. Все женщины также находились на третьем триместре беременности. Ни одна из женщин не рожала.Артериальное кровяное давление регистрировалось не менее двух раз с интервалом в 4 часа, когда пациент находился на постельном режиме от правой руки, с использованием стандартного сфигмоманометра с соответствующим размером манжеты одним и тем же врачом. Звук Короткова К5 (исчезновение всех звуков) использовали для определения диастолического артериального давления. Преэклампсия определяется минимальными критериями артериального давления ≥140 / 90 мм рт.ст. после 20 недель беременности и протеинурии 300 мг / 24 часа или тест-полоски 1+, согласно отчету Американской коллегии акушеров и гинекологов. Сила гипертонии при беременности [11].Контрольную группу составили 25 здоровых женщин с нормальным АД. 25 женщин из контрольной группы были отобраны таким же образом, как и пациенты из группы преэклампсии, в зависимости от возраста и гестационного возраста. Тридцать женщин, вошедших в группу преэклампсии, были дополнительно разделены на категории пациентов с преэклампсией тяжелой и легкой степени, каждая группа состояла из пятнадцати пациентов в соответствии с критериями, определенными в том же отчете, такими как систолическое артериальное давление 160 мм рт.ст. или выше или диастолическое артериальное давление. 110 мм рт. ст. или выше в двух случаях с интервалом не менее 4 часов, когда пациент находится в постельном режиме (если до этого времени не начата антигипертензивная терапия) Тромбоцитопения (количество тромбоцитов менее 100000 / микролитр) Нарушение функции печени, на что указывает аномально повышенный уровень крови концентрации печеночных ферментов (в два раза выше нормы), сильная стойкая боль в правом верхнем квадранте или эпигастральная боль, не поддающаяся лечению и не учитываемая альтернативными диагнозами, или и то и другое Прогрессирующая почечная недостаточность (концентрация креатинина в сыворотке выше 1.1 мг / дл или удвоение концентрации креатинина в сыворотке крови при отсутствии других почечных заболеваний) Отек легких Новые церебральные нарушения или нарушения зрения Один образец венозной крови (10 мл) был взят у пациентов с преэклампсией и здоровых контрольных в пробирки, содержащие этилендиаминтетрауксусную кислоту (K 2 EDTA). Образцы крови пациентов с преэклампсией были собраны до того, как они получили какие-либо лекарства. Сразу после отбора пробы плазмы отделяли центрифугированием в течение 15 мин при 4 ° C и замораживали при -80 ° C до анализа.Уровни N-концевого натрийуретического пептида про-C-типа в этих образцах плазмы измеряли методом ELISA с использованием набора EIA для N-концевого натрийуретического пептида про-C-типа (Biomedica, Австрия). Нижний предел обнаружения составил 0,55 пмоль / л. Изменения внутри анализа составляли 5–6% для диапазона концентраций 5,1–20 пмоль / л, а вариации между анализами составляли 1% для диапазона концентраций 5–20 пмоль / л, как указано производителем. Все образцы были проанализированы в двух экземплярах в одном и том же анализе. Сотрудники лаборатории не знали о клиническом состоянии женщин в исследуемых группах. В этом исследовании данные были представлены как средние значения ± стандартное отклонение и медианы (IQR). Для оценки данных использовались повторные измерения одностороннего дисперсионного анализа (ANOVA), критериев Краскела – Уоллиса и Манна – Уитни U , в которых значимое различие между группами определялось как. Тест множественных сравнений Данна, использованный в качестве апостериорного теста для получения значимых результатов. Анализ мощности проводился исходя из предположения, что в каждой группе имеется 14 пациентов со стандартным отклонением 4.Даже в этом сценарии рассчитанная мощность составила 0,87. Поскольку в каждой исследуемой группе у нас есть не менее 15 пациентов, мощность этого исследования кажется достаточной. Протокол нашего исследования был одобрен наблюдательным советом учреждения. Как показано в Таблице 1, не было значительных различий между группами исследования с нормотензивным, легким и тяжелым преэклампсией с точки зрения возраста матери, беременности, паритета и индекса массы тела (ИМТ). Кроме того, сроки беременности не различались между этими группами.У пациентов с тяжелой преэклампсией средний вес при рождении был значительно ниже, чем у пациентов с нормальным АД и пациентов с легкой преэклампсией () (Таблица 1). Различие в измерениях систолического и диастолического артериального давления было значительным между всеми тремя группами, и самое высокое давление наблюдалось в группе с тяжелой преэклампсией. Что касается биохимических данных, представленных в таблице 2, ожидаемая значительная разница в протеинурии среди всех трех групп. Наблюдалась значительная разница между группой с нормальным АД по сравнению с двумя другими исследуемыми группами. У пациентов с преэклампсией как легкой, так и тяжелой степени были более высокие уровни лактатдегидрогеназы (ЛДГ), чем у женщин с нормальным АД. Не было значительных различий в значениях аспартаттрансаминазы (AST) и аланинтрансаминазы (ALT) среди трех исследуемых групп.Уровни в плазме N-концевого натрийуретического пептида про-C-типа были значительно повышены в группе исследования тяжелой преэклампсии, отличаясь как от групп исследования с нормотензивной (), так и с легкой преэклампсией (), тогда как не было существенной разницы между легкой преэклампсией и нормотензивные учебные группы (). Средние значения (IQR) N-концевого натрийуретического пептида про-C-типа составили 6,48 (3,33) в группе женщин с нормальным АД, 7,37 (3,43) у пациентов с легкой преэклампсией и 11,52 (6.13) у пациентов с тяжелой преэклампсией. Распределение данных между исследуемыми группами и контрольной группой показано на рисунке 1. Анализ кривой оператора приемника (ROC) показал, что 10,17 пмоль / л является пороговым значением для тяжелой преэклампсии с LR 6,67, чувствительностью 80% (CI 51,9–95,7) и специфичность 88% (ДИ 68,8–97,5) (рис. 2). Натрийуретический пептид С-типа был третьим обнаруженным членом семейства натрийуретических пептидов.Семейство натрийуретических пептидов оказывает важное влияние на контроль артериального давления, объема и функцию почек [9]. Натрийуретический пептид С-типа, наиболее широко экспрессируемый натрийуретический пептид, по-видимому, является одним из наиболее важных членов этого семейства в регуляции кровяного давления. В японском исследовании было показано, что полиморфизм в трех первичных нетранслируемых областях (3′-UTR) гена натрийуретического пептида С-типа ( Nppc ) связан с гипертонией [12].Было обнаружено, что аллельные варианты в трех первичных нетранслируемых областях рецепторов натрийуретических пептидов A и C связаны с семейной гипертензией и повышенным систолическим артериальным давлением [13]. Кроме того, введение экзогенного натрийуретического пептида С-типа вызывает резкое падение артериального давления у людей, а также у некоторых других видов, таких как приматы и собаки [2]. Натрийуретический пептид С-типа также играет решающую роль в хронической регуляции артериального давления путем подавления симпатического оттока [14] и адренокортикотропного гормона [15], альдостерона [15] и синтеза вазопрессина [16].Преэклампсия — это синдром, поражающий почти все системы органов и характеризующийся гипертонией и протеинурией; однако изменения в уровнях натрийуретического пептида С-типа в отношении преэклампсии еще не ясны. Насколько нам известно, в литературе есть две публикации, в которых сравниваются уровни натрийуретического пептида С-типа у пациентов с преэклампсией и здоровых беременных женщин с нормальным давлением. Уровни натрийуретического пептида С-типа у пациентов с преэклампсией впервые были исследованы Stepan et al.В этом исследовании концентрацию натрийуретического пептида С-типа в плазме измеряли у пациентов с преэклампсией, беременных женщин с гестационной гипертензией и здоровых беременных. Они не смогли показать существенных различий между этими группами [9]. Второе исследование, проведенное Prickett et al. исследовали плазменные концентрации N-концевого натрийуретического пептида про-C-типа у беременных с преэклампсией и нормотензией во время родов. Поскольку натрийуретический пептид C-типа является пептидом местного действия с очень коротким периодом полужизни в кровотоке, они использовали пептид-предшественник с более длительным периодом полужизни натрийуретического пептида C-типа, который является N-концевым про-C-типом. натрийуретический пептид.В исследовании Prickett было обследовано 23 пациента, 13 из которых был поставлен диагноз преэклампсии без дискриминации в зависимости от тяжести синдрома, и они обнаружили значительные различия между двумя группами [10]. В нашем исследовании мы сравнили концентрацию N-концевого натрийуретического пептида про-С-типа в плазме у 55 субъектов, в том числе 25 здоровых беременных с нормальным АД и 30 беременных женщин с преэклампсией, все из которых находились в возрасте 30-40 недель. гестационный возраст. Исследовательская группа преэклампсии была далее разделена на группы преэклампсии с тяжелой и легкой степенью преэклампсии, каждая из которых состояла из 15 пациенток, разделенных в соответствии с критериями, определенными в отчете Целевой группы Американской коллегии акушеров и гинекологов по гипертонии при беременности [11] .Мы обнаружили, что концентрации в плазме N-концевого натрийуретического пептида про-C-типа были значительно выше в группе исследования с тяжелым преэклампсией по сравнению с группами исследования с умеренным преэклампсией и нормотензией, но между последними группами не было значительной разницы. Кроме того, средние концентрации в плазме N-концевого натрийуретического пептида про-C-типа во всех трех группах были ниже, чем концентрации, ранее отмеченные Prickett et al. Это могло произойти из-за того, что забор крови проводился во время сдачи крови в этом исследовании.Ранее было показано, что во время родов уровень натрийуретического пептида С-типа значительно повышается [17]. Кроме того, в исследовании Prickett et al. Было показано, что средние уровни N-концевого натрийуретического пептида про-C-типа были значительно ниже у беременных женщин по сравнению с небеременными женщинами, и эти уровни падают на протяжении всей беременности и остаются ниже чем у небеременных женщин того же возраста, по крайней мере до 36 недель. Эти значения более согласуются с нашими данными [18]. Наши данные показывают, что тяжесть преэклампсии может быть связана с циркулирующими уровнями N-концевого натрийуретического пептида про-С-типа.Espiner et al. показали, что преэклампсия и GHT с SGA были связаны с повышенным уровнем NTproCNP в материнской плазме [19]. Поскольку эндотелиальная дисфункция является одним из наиболее важных механизмов как в патофизиологии преэклампсии, так и в высвобождении эндотелием N-концевого натрийуретического пептида про-C-типа, эта конечная точка кажется логичной. Несмотря на то, что эти результаты заслуживают внимания, неясно, могут ли повышенные уровни N-концевого натрийуретического пептида про-C-типа быть полезными для прогнозирования или выявления тяжелой преэклампсии при клиническом применении.Одна из причин этого заключается в том, что повышение уровней в плазме N-концевого натрийуретического пептида про-C-типа, вероятно, является результатом эндотелиальной дисфункции, которая также усугубляет преэклампсию. Проспективно разработанное исследование может дать ответ на вопрос, можем ли мы предсказать более тяжелые случаи преэклампсии, которые, скорее всего, потребуют дальнейшего лечения и госпитализации, отслеживая уровни N-концевого про-C-натрийуретического пептида. Примечательно, что появляются многочисленные новые исследования прогностической и терапевтической роли ANP и BNP, а также двух других членов семейства натрийуретических пептидов при сердечно-сосудистых заболеваниях.Клинические испытания отметили преимущества и риски использования синтетического натрийуретического пептида A-типа (анаритида) и натрийуретического пептида B-типа (несиритида) для лечения артериальной гипертензии, сердечной недостаточности и почечной недостаточности [20]. Поскольку мы знаем, что инфузия натрийуретического пептида С-типа снижает артериальное давление, оно также может найти терапевтическое применение при лечении гипертонии у беременных женщин. Очевидно, что необходимы дальнейшие исследования для уточнения прогностической и терапевтической роли натрийуретического пептида С-типа в популяции пациентов с преэклампсией. Все данные, использованные для подтверждения результатов этого исследования, можно получить у соответствующего автора по запросу. Авторы заявляют, что у них нет конфликта интересов в связи с этой статьей. Ключевая мишень для терапевтических антител против тяжелого острого респираторного синдрома коронавируса 2 (SARS-CoV-2) представляет собой спайковый белок, тримерный белковый комплекс, каждый мономер которого содержит домен S1 и домен S2, которые опосредуют связывание с клетками-хозяевами и слияние мембран соответственно.Помимо рецепторсвязывающего домена (RBD), S1 имеет N-концевой домен (NTD). При поиске нейтрализующих антител основное внимание уделялось RBD. Chi et al. выделили антитела у 10 выздоравливающих пациентов и идентифицировали антитело, которое эффективно нейтрализует вирус, но не связывает RBD. Криоэлектронная микроскопия выявила эпитоп как NTD. Это антитело, нацеленное на NTD, может быть полезно в сочетании с антителами, нацеленными на RBD, в терапевтических коктейлях. Наука , в этом выпуске стр.650 При разработке терапевтических средств против тяжелого острого респираторного синдрома коронавируса 2 (SARS-CoV-2) можно было руководствоваться распределением эпитопов не только в рецепторсвязывающем домене (RBD) белка Spike (S), но и через полный белок Spike (S). Мы выделили и охарактеризовали моноклональные антитела (mAb) от 10 выздоравливающих пациентов с COVID-19. Три mAb показали нейтрализующую активность против аутентичного SARS-CoV-2. Одно mAb, названное 4A8, проявляет высокую эффективность нейтрализации как против аутентичного, так и против псевдотипированного SARS-CoV-2, но не связывает RBD.Мы определили эпитоп 4A8 как N-концевой домен (NTD) белка S путем определения с помощью криоэлектронной микроскопии его структуры в комплексе с S-белком с общим разрешением 3,1 ангстрем и локальным разрешением 3,3 ангстрем для белка 4A8. -NTD интерфейс. Это указывает на то, что NTD является многообещающей мишенью для терапевтических mAb против COVID-19. Глобальная вспышка COVID-19 представляет собой серьезную угрозу для здоровья человека ( 1 — 3 ). COVID-19 вызывается новым коронавирусом, коронавирусом 2 тяжелого острого респираторного синдрома (SARS-CoV-2), который представляет собой оболочечный вирус с положительной цепью РНК, вызывающий такие симптомы, как кашель, головная боль, одышка, миалгия, лихорадка и т. Д. и тяжелая пневмония у людей ( 1 , 3 — 5 ). SARS-CoV-2 является представителем рода β-коронавирусов, который также содержит SARS-CoV и MERS-CoV, вызвавшие эпидемии в 2002 и 2012 годах соответственно ( 6 , 7 ). SARS-CoV-2 имеет примерно 80% идентичность последовательности с SARS-CoV и использует тот же клеточный рецептор, ангиотензин-превращающий фермент 2 (ACE2) ( 8 — 16 ). Тримерный белок S украшает поверхность коронавируса и играет ключевую роль при проникновении вируса ( 17 , 18 ).Во время инфекции S-белок расщепляется на N-концевую субъединицу S1 и C-концевую субъединицу S2 протеазами хозяина, такими как TMPRSS2 ( 18 , 19 ), и изменяет конформацию от состояния до слияния к состоянию после слияния ( 20 ). ). S1 и S2 содержат внеклеточный домен (ECD; от 1 до 1208 аминокислот) и единственную трансмембранную спираль и опосредуют связывание рецептора и слияние мембран, соответственно ( 16 ). S1, который состоит из N-концевого домена (NTD) и рецепторсвязывающего домена (RBD), имеет решающее значение для определения тканевого тропизма и диапазонов хозяев ( 21 , 22 ).RBD отвечает за связывание с ACE2, тогда как функция NTD не совсем понятна. В некоторых коронавирусах NTD может распознавать специфические сахарные фрагменты при начальном прикреплении и может играть важную роль в переходе S-белка от пре-слияния к пост-слиянию ( 23 — 26 ). NTD белка S БВРС-КоВ может служить критическим эпитопом для нейтрализации антител ( 26 ). Моноклональные антитела (mAb) SARS-CoV-2 S, нацеленные на белок, с мощной нейтрализующей активностью, являются основным направлением разработки терапевтических вмешательств для COVID-19 ( 27 — 29 ).Во многих исследованиях сообщалось о функциях и структурах нейтрализующих SARS-CoV-2 антител, которые нацелены на RBD и ингибируют ассоциацию между S-белком и ACE2 ( 28 — 34 ). Антитела, нацеленные на RBD, применяемые индивидуально, могут вызывать мутации устойчивости в вирусе ( 26 ). Антитела, которые нацелены на эпитопы, не относящиеся к RBD, могут быть добавлены в терапевтические коктейли антител для SARS-CoV-2. Таким образом, мы стремились идентифицировать антитела к различным областям белка S и к белку нуклеокапсида (N). Для выделения mAb и анализа гуморальных антител к SARS-CoV-2 мы собрали плазму и мононуклеарные клетки периферической крови (PBMC) у 10 китайских пациентов. который вылечился от инфекции SARS-CoV-2. Возраст доноров от 25 до 53 лет. Интервал от даты подтверждения заболевания до даты сбора крови составлял от 23 до 29 дней для пациентов с 1 по 5 и от 10 до 15 дней для пациентов с 6 по 10 (таблица S1).Мы оценили титры связывающих антител в плазме с различными фрагментами белка SARS-CoV-2 S, включая полный ECD, S1, S2 и RBD, а также с белком N. Плазма всех пациентов, кроме донора 2, связывалась со всеми пятью белковыми сегментами SARS-CoV-2, тогда как плазма донора 2 распознавала только S-ECD и S2 (рис. 1A). Нейтрализующие способности плазмы против аутентичного SARS-CoV-2 и псевдотипированного SARS-CoV-2 с вектором ВИЧ коррелированы [коэффициент корреляции ( r ) = 0,6868, P <0.05] (рис. 1Б). Эти результаты показывают, что гуморальные иммунные ответы были специфически вызваны у всех 10 пациентов во время их естественного заражения SARS-CoV-2. ( A ) Реакции плазмы на белки SARS-CoV-2. S-ECD (внеклеточный домен S-белка), S1, S2, RBD (рецептор-связывающий домен) и N-белок использовали в ELISA для проверки связывания плазмы. Плазма здоровых доноров была использована в качестве контроля, и пороговые значения были рассчитаны как оптическая плотность (OD) 450 контроля × 2.1. Данные были показаны со средним значением и стандартным отклонением репрезентативного эксперимента. ( B ) Корреляция между титрами аутентичных нейтрализующих антител (NAb) SARS-CoV-2 и псевдотипными титрами NAb SARS-CoV-2 в плазме. Анализы нейтрализации плазмы против аутентичного SARS-CoV-2 проводили с использованием клеток Vero E6, а нейтрализацию против псевдотипированного SARS-CoV-2 определяли с использованием клеток ACE2-293T. Корреляции рассчитывались с помощью теста корреляции Пирсона в Graphpad 7.0. ( C ) Сортировка методом проточной цитометрии из PBMC 10 выздоравливающих пациентов.( D ) Распределение семейств генов V в тяжелой и легкой цепях всех отдельных клонов (общее количество показано в центре круговых диаграмм) для каждого донора. ( E ) Показано количество аминокислотных (AA) и общих нуклеотидных (Nt) мутаций в зародышевой линии всех клональных последовательностей, идентифицированных в (D). ( F ) Аминокислотные длины CDR3 VH и VL всех клональных последовательностей, идентифицированных в (D). Чтобы изолировать S-протеин-специфические mAb, мы сначала отсортировали иммуноглобулин G-положительные (IgG + ) В-клетки памяти из PBMC выздоравливающих пациентов 1–5 с помощью проточной цитометрии, используя S-ECD в качестве зонда (рис.1С). Процент S-ECD-реактивных В-клеток IgG + колеблется от 0,56 до 11%, что выявлено с помощью сортировки активирующих флуоресценцию клеток (FACS). Чтобы избежать потери B-клеток с низкими копиями S-ECD-специфических рецепторов на поверхности клеток, мы отсортировали плазматические B-клетки из смешанных PBMC, полученных от других пяти выздоравливающих пациентов (пациенты с 6 по 10), без использования белка S-ECD в качестве зонда в потоке. цитометрия. Процент В-клеток плазмы в В-клетках CD3-CD19 + составил 12,8%, что выше, чем процент В-клеток памяти в В-клетках CD3-CD19 + (рис.1С). Из отсортированных B-клеток мы идентифицировали 9, 286, 43, 12 и 26 клонов одиночных B-клеток от пациентов от 1 до 5, соответственно, и 23 клона одиночных B-клеток из смешанных PBMC пациентов от 6 до 10 ( Рис. 1D). Распределение семейств секвенированных генов тяжелой цепи (IgH) было сопоставимым среди 10 доноров, причем Vh4 был наиболее часто используемым геном VH, тогда как разные доноры демонстрировали различные предпочтения в отношении семейств генов легкой цепи (IgL) (рис. 1D). . Комбинация V3 и J4, V3 и D3, а также D3 и J4 была наиболее частым использованием для семейства генов IgH (рис.S1). Среднее количество мутаций аминокислот на mAb из В-клеток памяти варьировалось от 17,50 до 48,04 для доноров 1-5, соответственно, тогда как mAb из B-клеток плазмы обладали в среднем 13,99 аминокислотных мутаций для доноров от 6 до 10 (фиг. 1E). Человеческие антитела, вызванные многократным воздействием различных антигенов, вызывают в среднем 26,46 аминокислотных мутаций на Ab, как сообщалось ранее ( 35 ). Эти результаты показывают, что естественная инфекция SARS-CoV-2 вызвала высокий уровень соматической гипермутации (SHM) в B-клетках памяти.Длина определяющей комплементарность области 3 (CDR3) для антител была сходной у доноров, при этом средняя длина этих CDR3 составляла от 13,9 до 17,7 для VH и от 9,3 до 10,1 для VL (фиг. 1F). Длина CDR3 этих mAb была больше, чем у антиген-специфических иммунных рецепторов (среднее значение 12,7 для VH и 6,5 для VL, соответственно), о которых сообщалось ранее ( 36 ). Для скрининга на S-белок-специфические антитела мы определили специфичность связывания с использованием иммуноферментного анализа (ELISA) для 399 человеческих mAb, отсортированных выше.От доноров 1-5, соответственно, были идентифицированы 1, 16, 1, 3 и 9 антител, специфичных к S-ECD. Всего было идентифицировано 35 S-ECD-специфических mAb от доноров от 6 до 10 (рис. 2A). Мы дополнительно охарактеризовали доменную специфичность 35 mAb с различными фрагментами белка S, включая S1, S2 и RBD (рис. 2A). S-реактивные mAb классифицируются на четыре основные группы на основе значений их средней эффективной концентрации (EC 50 ) (фиг. 2A). Группа 1 распознает только S-ECD. Группа 2 распознает S-ECD и S1, при этом подгруппа 2A связывает S-ECD и S1, а подгруппа 2B связывает S-ECD, S1 и RBD.Группа 3 взаимодействует как с S1, так и с S2, где подгруппа 3A нацелена на RBD, а подгруппа 3B не может связать RBD. Группа 4 распознает S-ECD и S2. Только четыре mAb распознают RBD среди 35 S-специфичных mAb (рис. 2, A и B). ( A ) Тепловая карта, показывающая связывание mAb с различными типами белков-шипов, определенное с помощью ELISA. Показано значение EC 50 для каждой комбинации S-mAb с темно-красным, оранжевым, желтым или белым затенением, указывающим на высокое, промежуточное, низкое или отсутствие детектируемого связывания соответственно.EC 50 указаны значения более 10 000 нг / мл (>). ( B ) Кривые связывания репрезентативных mAb. CR3022 является контролем, который, как сообщается, связывает SARS-CoV и SARS-CoV-2 RBD. Данные были показаны со средним значением и стандартным отклонением репрезентативного эксперимента. ( C ) Тепловая карта, показывающая конкурирующее связывание некоторых репрезентативных S-реактивных mAb, проанализированных с помощью ELISA. Числа в рамке указывают процент связывания детектирующего mAb в присутствии блокирующего антитела по сравнению со связыванием детектирующего mAb в отсутствие блокирующего антитела.MAb считали конкурирующими, если процент ингибирования составлял <30% (черные прямоугольники с белыми числами). Было установлено, что mAb не конкурируют за один и тот же сайт, если процент составляет> 70% (белые квадраты с красными числами). Серые прямоугольники с черными числами указывают на промежуточный фенотип (от 30 до ~ 70%). ( D ) Филогенетические деревья всех S-специфичных mAb. Мы провели анализ конкурентного связывания с использованием ELISA для нескольких репрезентативных mAb, чтобы определить, есть ли перекрывающиеся антигенные сайты между разными mAb, с CR3022, используемым в качестве mAb положительного контроля, которое сообщило о связывании SARS-CoV-2 RBD (рис.2С) ( 37 ). Среди этих mAb 4A8 в группе 2A конкурировали с 1M-1D2 в группе 2B. Другое RBD-реактивное mAb, 2M-10B11 в группе 2B, конкурировало с CR3022, предполагая перекрывающиеся эпитопы на RBD для этих двух mAb. Эти результаты показывают, что ответы антител, вызванные естественной инфекцией SARS-CoV-2, различались по распознаванию эпитопов S-белков. Чтобы охарактеризовать разнообразие в использовании генов и созревании аффинности, филогенетические деревья этих S-ECD-специфичных mAb были проанализированы на основе аминокислотных последовательностей VHDJH и VLJL с использованием метода объединения соседей в программном обеспечении MEGA7 ( 38 ).Результаты показывают, что использование гена VH очень разнообразно среди 35 mAb от 10 доноров, причем Vh4-30 является наиболее часто используемым геном зародышевой линии. Среди S1, S2 или RBD-реактивных mAb не было выявлено особо предпочтительного гена VH (фиг. 2D). Процент идентичности последовательностей вариабельного гена тяжелой цепи варьировал от 40,9 до 97,6% в 35 S-ECD-специфических mAb (рис. S2 и таблица S2). Сначала мы провели исследования нейтрализации in vitro 35 S-ECD-специфичных mAb с использованием аутентичных SARS-CoV-2 в клетках Vero-E6 (рис.3А). Из 35 антител, специфичных для S-ECD, только три антитела нейтрализовали подлинный SARS-CoV-2. MAb 1M-1D2, 4A8 и 0304-3h4 проявляли нейтрализующую способность от средней до высокой с ЕС 50 28, 0,61 и 0,04 мкг / мл соответственно. Как и ожидалось, контрольное mAb, нацеленное на RBD, CR3022, не смогло нейтрализовать подлинный SARS-CoV-2 ( 37 ). Более того, хотя конкурирующее с CR3022 mAb, 2M-10B11, связывалось с RBD SARS-CoV-2 с EC 50 , равным 5 нг / мл (рис. 2A), оно также не смогло нейтрализовать подлинный SARS-CoV-2. .Эти результаты предполагают, что аффинность связывания mAb с RBD не полностью коррелирует с нейтрализующей способностью mAb. Для дальнейшего изучения ингибирующей активности трех аутентичных SARS-CoV-2 — нейтрализующих mAb — 4A8, 0304-3h4 и 1M-1D2 — мы протестировали РНК-нагрузку аутентичного SARS-CoV-2 в клетках Vero-E6, обработанных каждым из них. mAb с использованием количественной полимеразной цепной реакции (ПЦР) в реальном времени (фиг. 3B). В соответствии с результатами анализа цитопатического эффекта (ЦПЭ) (фиг. 3A), mAb 0304-3h4 и 4A8 показали более высокую ингибирующую способность, чем 1M-1D2 (фиг.3Б). ( A ) Нейтрализация S-реактивных mAb до аутентичного SARS-CoV-2 в клетках Vero-E6. ( B ) Подлинную нагрузку РНК вируса SARS-CoV-2 определяли в клетках Vero-E6, обработанных S-реактивными mAb, с помощью количественной ПЦР. Процент инфицирования рассчитывали как отношение нагрузки РНК в лунках, обработанных mAb, к нагрузке в лунках, содержащих только вирус. ( C ) Нейтрализация S-реактивных mAb против ВИЧ-инфицированного псевдотипированного SARS-CoV-2 в клетках ACE2-293T.Данные представлены как среднее ± стандартное отклонение репрезентативного эксперимента. Затем мы провели анализы репортерного гена люциферазы для всех 35 S-связывающих mAb с использованием ВИЧ-векторных псевдотипированных SARS-CoV-2 ( 39 ), среди которых три mAb проявляли нейтрализующую активность против псевдотипированного вируса (фиг. 3C). 4A8 защищал клетки ACE2-293T с EC 50 49 мкг / мл. Хотя mAb 2M-10B11 и 9A1 не нейтрализовали аутентичный SARS-CoV-2, 2M-10B11 защищал от псевдотипированного вируса с помощью EC 50 170 мкг / мл, а 9A1 обеспечивал слабую защиту.К нашему удивлению, нейтрализации 0304-3h4 и 1M-1D2 не наблюдалось (рис. 3C). Несоответствие между результатами для псевдотипированного SARS-CoV-2 по сравнению с аутентичным SARS-CoV-2 также наблюдалось для mAb против MERS-CoV ( 40 , 41 ) и может быть вызвано различным представлением полученного S-белка. от различных факторов окружающей среды, которым подверглись вирусы, таких как клетки, используемые для анализов нейтрализации или для получения псевдотипированных или аутентичных вирионов ( 42 ).На основе этих результатов 4A8 является потенциальным кандидатом для лечения SARS-CoV-2, поскольку он продемонстрировал сильную нейтрализующую способность против как подлинного, так и псевдотипного SARS-CoV-2. Чтобы определить возможный механизм нейтрализации mAb, мы определили аффинности связывания пяти mAb с потенциальной нейтрализующей активностью в отношении различных сегментов белка S, включая полный S-ECD и домены S1, S2 и RBD — с использованием интерферометрии биослоя (BLI).Все пять протестированных mAb связались с S-ECD с высоким сродством; константы равновесной диссоциации ( K d ) были менее 2,14 нМ (фиг. 4A). 4A8 и 1M-1D2 связаны с S1 с K d 92,7 и 108 нМ соответственно, тогда как 0304-3h4 и 9A1 нацелены на S2 с K d 4,52 и <0,001 нМ, соответственно (рис. 4A , Нижний). Более того, 2M-10B11 связывает RBD с K d из 24,3 нМ, что было получено с использованием модели гетерогенного лиганда из-за эффектов авидности (рис.4А, внизу). ( A ) Сенсограммы BLI и кинетика связывания mAb с S-белками. Глобальные кривые подгонки показаны черными линиями. K d вычисляли с использованием модели связывания 1: 1 в Data Analysis Software 9.0, за исключением 2M-10B11, в котором использовалась модель гетерогенного лиганда из-за эффекта авидности. ( B ) Связывание белка S со сверхэкспрессирующими ACE2 человеческими клетками 293T определяли с помощью проточной цитометрии.После предварительной инкубации белка S с каждым указанным mAb смеси mAb-S добавляли к клеткам, экспрессирующим ACE2. Клетки окрашивали флуоресцеинизотиоцианатом против человеческого IgG (связывание mAb, ось x ) и анти-His (связывание S, ось y ). Показаны проценты дважды положительных клеток. Ранее сообщалось, что контрольные mAb CR3022 и 1A8 связывают SARS-CoV RBD и гликопротеин Марбург, соответственно, и белок ACE2-Fc представлял собой белок ACE2 человека, конъюгированный с Fc человека. Чтобы исследовать, блокируют ли эти mAb связывание S-белка с ACE2, мы провели проточную цитометрию с использованием клеток 293T эмбриональной почки человека (HEK), экспрессирующих человеческий ACE2. Как и ожидалось, только 2M-10B11 из пяти mAb и ACE2-Fc предотвращали связывание белка S с ACE2. В присутствии 2M-10B11 только 0,52% клеток были дважды положительными по IgG и S-белку (фиг. 4B). CR3022, который конкурирует с 2M-10B11, не блокирует связывание S с ACE2. Контрольное mAb 1A8, нацеленное на гликопротеин Марбург, также не препятствовало связыванию, а 5.13% двойных положительных результатов могут быть связаны с неспецифическим связыванием 1A8 с белком S. 4A8 также не препятствовал связыванию белка S с ACE2. mAb 4A8 было сверхэкспрессировано и очищено смолой с протеином A, а S-ECD SARS-CoV-2 был очищен с помощью аффинной смолы M2 и эксклюзионной хроматографии (SEC). Белок 4A8 и S-ECD смешивали и инкубировали при стехиометрическом соотношении ~ 1,2: 1 в течение 1 часа и наносили на SEC для удаления избыточных белков (рис.S3A). Фракцию, содержащую комплекс, концентрировали для приготовления образцов для криоэлектронной микроскопии (крио-ЭМ). Чтобы исследовать взаимодействия между 4A8 и S-белком, мы решили крио-ЭМ структуру комплекса с общим разрешением 3,1 Å (рис. 5 и фильм S1). Подробности подготовки крио-ЭМ проб, сбора и обработки данных, а также построения модели можно найти в дополнительных материалах, материалах и методах (рис. S3 — S5). Белок S демонстрирует асимметричные конформации, аналогичные ранее описанным структурам ( 21 , 22 ), причем один из трех RBD находится в «верхней» конформации, а два других RBD — в «нижней» конформации (рис.5). КриоЭМ-карта комплекса с цветными доменами показана слева, а два перпендикулярных вида всей структуры показаны справа. Тяжелая и легкая цепи 4A8 окрашены в синий и пурпурный цвета соответственно. NTD тримерного S-белка окрашены в оранжевый цвет. Один «верхний» RBD и два «нижних» RBD тримерного S-белка окрашены в зеленый и голубой цвета соответственно. В комплексе S-белок-4A8 каждый тримерный S-белок связан с тремя растворенными Fab-фрагментами 4A8, каждый из которых взаимодействует с одним NTD-белком S.Несмотря на разные конформации трех протомеров белка S, интерфейс между 4A8 и каждым NTD идентичен (рис. 5 и рис. S3I). Качество карты в области NTD-4A8 было улучшено за счет целенаправленного уточнения до локального разрешения 3,3 Å, что позволяет надежно анализировать взаимодействия между NTD и 4A8. Ассоциация с 4A8, по-видимому, стабилизирует эпитоп NTD, который не виден только в описанной структуре белка S ( 21 , 22 ). Благодаря высокому разрешению NTD мы смогли построить структурную модель для пяти новых петель для NTD, обозначенных N1 (остатки с 14 по 26), N2 (остатки с 67 по 79), N3 (остатки с 141 по 156), N4 ( остатки 177–186) и N5 (остатки 246–260), среди которых петли N3 и N5 опосредуют взаимодействие с 4A8 (рис.S5A). Кроме того, в этой структуре идентифицированы три новых сайта гликозилирования (Asn 17 , Asn 61 и Asn 149 ) на NTD (рис. S6). Тяжелая цепь 4A8 в основном участвует в связывании с NTD главным образом через три определяющие комплементарность области (CDR), названные CDR1 (остатки 25-32), CDR2 (остатки 51-58) и CDR3 (остатки 100-116) (Рис. 6A и рис. S5B). Интерфейс состоит из разветвленной сети гидрофильных взаимодействий, а площадь скрытой поверхности на границе раздела 4A8-NTD составляет 832 Å 2 .Arg 246 на петле N5 NTD представляет собой один сайт стыковки, который стабилизируется Trp 258 , одновременно взаимодействуя с Tyr 27 и Glu 31 из 4A8 на CDR1 (фиг. 6B). На петле N3 NTD Lys , 150, и Lys , 147, , соответственно, образуют солевые мостики с Glu 54 и Glu 72 из 4A8 (фиг. 6C). Lys 150 также связан водородом (H) с 4A8-Tyr 111 , тогда как His 146 образует водородную связь с 4A8-Thr 30 (рис.6С). Помимо гидрофильных взаимодействий, Trp 152 и Tyr 145 на петле N3 NTD также взаимодействуют с Val 102 , Pro 106 и Phe 109 на CDR3 4A8 посредством гидрофобных и / или π-π взаимодействия (рис. 6D). Кроме того, сайт гликозилирования Asn 149 на NTD близок к интерфейсу 4A8-NTD, из которых N-гликаны могут участвовать во взаимодействиях на интерфейсе (фиг. 6A и фиг. S6). ( A ) Обширные гидрофильные взаимодействия на границе раздела между NTD и 4A8. Показан только один NTD-4A8. ( B — D ) Подробный анализ интерфейса между NTD и 4A8. Полярные взаимодействия обозначены красными пунктирными линиями. Остатки, участвующие в гидрофобных взаимодействиях, представлены сферами. Существует острая необходимость в профилактических и терапевтических вмешательствах в отношении инфекций SARS-CoV-2, учитывая продолжающуюся пандемию COVID-19.Наша работа показывает, что природные моноклональные антитела SARS-CoV-2 человека, выделенные из В-клеток 10 восстановленных доноров, различаются по использованию генов и распознаванию эпитопа S-белка. Большинство выделенных mAb не распознают RBD, и все mAb, нейтрализующие аутентичный SARS-CoV-2, не могут ингибировать связывание белка S с ACE2. Эти неожиданные результаты предполагают наличие других важных механизмов нейтрализации SARS-CoV-2 в дополнение к подавлению взаимодействия вируса с рецептором. Нацеленные на S1 mAb 4A8 не блокируют взаимодействие между ACE2 и S-белком, но демонстрируют высокие уровни нейтрализации как против аутентичного, так и против псевдотипированного SARS-CoV-2 in vitro. Сообщалось, что многие нейтрализующие антитела против SARS-CoV-2 нацелены на RBD белка S и блокируют связывание между RBD и ACE2 ( 28 — 30 , 32 — 34 ). Наши результаты показывают, что 4A8 связывается с NTD белка S с сильной нейтрализующей активностью.Предыдущее исследование показало, что mAb 7D10 может связываться с NTD S-белка MERS-CoV, вероятно, путем ингибирования связывания RBD-DPP4 и конформационного изменения S-белка перед слиянием до пост-слияния ( 26 ). Мы выровняли кристаллическую структуру 7D10 в комплексе с NTD белка S MERS-CoV с нашей сложной структурой и обнаружили, что границы раздела между mAb и NTD частично перекрываются (рис. S7). 7D10 может ингибировать взаимодействие между MERS-CoV и DPP4 через свою легкую цепь, которая близка к RBD.В нашем комплексе легкая цепь 4A8 находится далеко от RBD (рис. S7). Поэтому мы предполагаем, что 4A8 может нейтрализовать SARS-CoV-2, сдерживая конформационные изменения белка S. Кроме того, выравнивание последовательностей белков S из SARS-CoV-2, SARS-CoV и MERS-CoV выявило различные поверхностные последовательности NTD, которые соответственно распознаются разными mAb (рис. S8). В этой работе сообщается о полностью нейтрализующем человеческом mAb, распознающем уязвимый эпитоп NTD на S-белке SARS-CoV-2, функционирующем с механизмом, который не зависит от ингибирования связывания рецептора.Комбинация 4A8 с антителами, нацеленными на RBD, может избежать ускользающих мутаций вируса и служить многообещающим «коктейльным» терапевтическим средством. Информация, полученная в результате этих исследований, может быть использована для разработки дизайна вакцины на основе структуры против SARS-CoV-2. Доказательства in vivo.
Ацетилирование CobB
L не вызывает деградацию CobB L in vivo. Обсуждение
Может ли YiaC ацетилировать ε-аминогруппу боковых цепей лизина?
Заключительные замечания.
Материалы и методы
Доступность данных.
Благодарности
Сноски
Идентификация и концентрации в плазме N-концевого фрагмента предсердного натрийуретического фактора у человека * | The Journal of Clinical Endocrinology & Metabolism
Связаны ли уровни натрийуретических пептидов N-терминального типа Pro-C с тяжестью преэклампсии?
1. Введение
2. Материалы и методы
3. Результаты
Переменная Нормотензивные пациенты ( n = 25) Пациенты с легкой преэклампсией ( n = 15) Пациенты с тяжелой преэклампсией ( n ) Возраст матери (год) 27.9 ± 4,8 29,6 ± 3,8 26,7 ± 5,2 Gravida 3 (1–6) 2 (1–8) 3 (1–4) Четность 2 (0–4) 2 (0–6) 2 (1–3) Индекс массы тела матери (кг / м 2 ) 28,33 ± 3,8 26,21 ± 3,2 28,64 ± 2,8 Масса при рождении (грамм) 3371 ± 484 3106 ± 538 2656 ± 614 Гестационный возраст (неделя) 35.6 ± 1,9 35,5 ± 2,7 35,6 ± 1,7 Систолическое артериальное давление (мм рт. Ст.) 100 (10) 140 (10) 160 (10) Диастолическое артериальное давление (мм рт. ) 70 (20) 90 (2,5) 100 (7,5) Значения представлены как среднее ± стандартное отклонение и медианы (значения IQR). по сравнению с нормотензивными женщинами и пациентами с легкой преэклампсией.по сравнению с женщинами с нормальным давлением. Переменная Нормотензивные пациенты ( n = 25) Пациенты с легкой преэклампсией ( n = 15) Пациенты с тяжелой преэклампсией ( n ) Протеинурия (мг / 24 ч) — 613.8 ± 306,5 3842,5 ± 2122,9 † Креатинин плазмы (мг / дл) 0,5 (0,1) 0,6 (0,2) 0,7 (0,1) LDH (U / L) 151,6 (37) 219,6 (79,7) 277,6 (66,9) AST (U / L) 17,2 (11,8) 21,2 (8,4) 21,4 (12,8) ALT ( Ед / л) 11,6 (4,6) 12,3 (8,0) 12,9 (7,8) N-концевой натрийуретический пептид про-C-типа (пмоль / л) 6.48 (3,33) 7,37 (3,43) 11,52, (6,13) Значения представлены как среднее ± стандартное отклонение и медианы (значения IQR). по сравнению с женщинами с нормальным давлением. по сравнению с женщинами с легкой преэклампсией. по сравнению с женщинами с нормотензией и легкой преэклампсией.
4. Обсуждение
5. Заключение
Доступность данных
Конфликты интересов
Нейтрализующее человеческое антитело связывается с N-концевым доменом белка Spike SARS-CoV-2
Поражение SARS-CoV-2 на новом месте
Abstract
Результаты
Выделение mAb человека из B-клеток памяти и B-клеток плазмы
Профили связывания SARS-CoV-2 S-белок-специфичных человеческих mAb
Нейтрализующая активность SARS-CoV-2 S-специфических mAb человека
Характеристика связывания mAb-кандидатов
Крио-EM структура комплекса между 4A8 и S-ECD
Распознавание NTD с помощью 4A8
Обсуждение
1, 81 8015 и 31930059), Программой ключевых исследований и разработок провинции Чжэцзян. (2020C04001), чрезвычайный проект SARS-CoV-2 Департамента науки и технологий провинции Чжэцзян (2020C03129), Программа внедрения ведущей инновационной и предпринимательской группы Ханчжоу, национальный крупный проект в области науки и технологий Министерства науки и технологий of China, (2018ZX10101003-005-007), и Специальная программа исследований новой коронавирусной пневмонии Университета Вестлейк. Вклад авторов: W.C., Q.Z. и J.L. разработали проект. X.C., R.Y., Ju.Z., G.Z., Y.Z., Y.G., Y.L., L.X., M.H., Z.Z., P.F., Y.D., Z.C., Ji.Z., X.S., Y.C., L.F., L.H., J.X. и C.Y. сделал эксперименты. Все авторы внесли свой вклад в анализ данных. X.C., R.Y., J.L., Q.Z. и W.C. написал рукопись. Конкурирующие интересы: W.C., J.L., X.C., Ju.Z., L.F., C.Y., J.X., L.H., G.Z., P.F., M.H., Y.D., X.S., Y.C. и Ji.Z. указаны как изобретатели в находящейся на рассмотрении заявке на патент на mAb 4A8.Остальные авторы заявляют, что у них нет конкурирующих интересов. Доступность данных и материалов: Координаты атомов и карты плотности крио-ЭМ S-белка SARS-CoV-2 в комплексе, связанном с 4A8 (PDB: 7C2L; полная карта: EMD-30276, уточненная карта, ориентированная на интерфейс антитело-эпитоп : EMD-30277) были депонированы в Банк данных белков (www.